Кому мешает ДНК-генеалогия: Общий предок и мутации в гаплотипах потомков.

Начало...
Сколько всего имеется основных родов, или гаплогрупп человечества?
Гаплогруппы не просто соответствуют своим родам, но образуют определённую последовательность, лестницу гаплогрупп, показывающих их иерархию - последовательный, ступенчатый переход от точки расхождения африканских и неафриканских популяций (примерно 160 тыс. лет назад) до самой недавней гаплогруппы R, образовавшейся примерно 30 тыс. лет назад. Эта лестница называется филогенетическим деревом гаплогрупп и их снипов. Все гаплогруппы и субклады на дереве должны включать снипы «вышестоящих» гаплогрупп и субкладов. То есть принцип «лесенки» должен выполняться. Преемственность узловых родов человечества должна соблюдаться.
Всего в мире насчитывают 20 основных гаплогрупп, которые обозначают буквами латинского алфавита, от А до Т, хотя систему порой нарушают. В последнее время в классификацию добавили гаплогруппы A0 и A00, хотя их носителей обнаружили в количестве считанных единиц, все они живут в Африке. Но они настолько отстоят по снип-мутациям от всех остальных людей на Земле (тестированных на мутации в ДНК), что их пришлось выделить в отдельный род-гаплогруппу. Помимо этого, были идентифицированы промежуточные, сводные гаплогруппы, такие, как СТ, DE, GHIJK и другие, так что минимальный состав генеалогического дерева мужской половины человечества включает уже 39 основных гаплогрупп, то есть главных уровней Y-хромосомной генеалогической структуры. С подгруппами это составляет уже много сотен.
Если же считать все уровни генеалогического дерева (уровень в этом случае - это субклад), то гаплогруппа R1a, основная гаплогруппа (род) этнических русских, уже насчитывает 66 субкладов, гаплогруппа R1b - 443 субклада, и это при том, что на диаграмме ниже они обе входят в сводную гаплогруппу R, которой примерно 30 тысяч лет со времени образования. Образовалась гаплогруппа R при появлении необратимой мутации в виде спонтанного превращения одного нуклеотида в другой в ДНК (Y-хромосоме) патриарха, или его выжившего потомка, в свою очередь, потомки которого дожили до настоящего времени в количестве более миллиарда человек. Если точнее, то у него, патриарха гаплогруппы R, по сравнению с ДНК его отца (относящегося к гаплогруппе Р) произошло спонтанное превращение аденина в гуанин, и это произошло в участке Y-хромосомы под номером 15 миллионов 581 тысяч 983. Всего же в Y-хромосоме мужчин насчитывается, как сообщалось выше, примерно 58 миллионов нуклеотидов.
Можно ли рассчитать, когда жил общий предок всех сегодняшних людей на Земле?
Можно, с определенной степенью допущений. Первое и самое главное допущение - что общий предок сегодняшних людей на Земле на самом деле существовал. Естественно, речь не идет о том, что на Земле когда-то жил один мужчина, и с ним рядом была одна женщина, как повествует библейская притча. Такого никогда не было. Во все времена было множество мужчин и множество женщин, а до них было множество самцов и самок, предшественников тех самых мужчин и женщин, и переход одних в других занял миллионы лет, и никто не может дать четких определений, в чем выражался этот переход, это все вопросы определений, которых, в общем-то, нет. Да, наверное, никого это отсутствие четких определений особенно и не беспокоит.
Чисто концептуально, «общий предок» всех людей на сегодняшней планете - это тот, потомство которого выжило, в отличие от множества его современников, и продолжилось в потомках до настоящего времени. Помимо этого - это был тот человек, у которого было как минимум два сына, потомство которых выжило до настоящего времени. Как следствие первых двух условий, все потомки унаследовали снип-мутации своего «первопредка», и добавили снип-мутации в последующем. Поэтому есть еще одно условие, точнее, следствие - мутации в гаплотипах потомков при экстраполяции в прошлое сходятся к «первопредку». Таким образом, можно определить его гаплотип, независимо от того, как давно он жил.
Но в реальности картина более сложная - ДНК-линия от первопредка к сегодняшним потомкам часто выглядит как цепь сосисок с перетяжками во многих местах. Эти места - бутылочные горлышки популяции, и порой перетяжки образуют такие сужения, что при расчетах они тоже формально соответствуют одному человеку, патриарху после прохождения бутылочного горлышка. Иначе говоря, имеем последовательную цепь патриархов. Это легко представить на примере хрестоматийного «последнего из могикан» Фенимора Купера. Вспомним сюжет - от всего племени могикан остался один мужчина. Умрет он - и вся линия ДНК, идущая из глубин, возможно, 12-15 тысяч лет назад, оборвется. Для следующего поколения американских индейцев могикан как бы и не существовало, несмотря на их многотысячелетнюю историю. Но если «последний из могикан» выживет, женится, обзаведется сыновьями - он станет патриархом для последующих поколений его рода.
Подобных случаев в истории Земли было, видимо, неисчислимое количество. В итоге вопрос про «общего предка человечества» несколько расплывается. В каждой прямой ДНК-линии её общий предок жил относительно недавно - где сотни лет назад, где тысячу-другую лет назад. Но совокупность ДНК-линий, даже только выживших, помещает общих предков на 20-40 тысяч лет назад в каждой гаплогруппе. Совокупность ДНК-линий разных гаплогрупп помещает общего предка неафриканцев на 64±6 тысяч лет назад, до наиболее отдаленного бутылочного горлышка популяции, а африканцев - примерно на 217 тысяч лет назад, но это не потому, что африканские предки древнее, а потому что по какой-то причине совокупность африканских (по их нынешнему месту обитания) ДНК-линий прошла бутылочное горлышко популяции намного ранее.
Можно ли считать, что 217 тысяч лет назад жил общий предок современного человечества? Нет, нельзя. Потому что неафриканские и африканские ДНК-линии сходятся к общему предку примерно 160 тысяч лет назад. Именно тогда разошлись африканские и неафриканские (по нынешнему месту обитания) ДНК-линии. Но глубже во времени уходит общий эволюционный ствол африканцев и неафриканцев, который расходится на гаплогруппы А00 и А0-Т примерно 217 тысяч лет назад. У нас нет оснований не считать носителей гаплогрупп А00 и А0-Т не членами вида Homo sapiens, так что общий предок современного человечества уходит глубже 217 тысяч лет назад. Но еще глубже, 300-400-500 тысяч лет назад, уже времена образования неандертальцев, которых вряд ли можно считать общими предками современного человечества. Вот и очерчены рамки времен возникновения общего предка современного человечества. Называть его «Адам», чем увлекаются некоторые, нет никаких оснований.
Почему в ДНК-генеалогии за поколение принимается 25 лет? Ведь протяженность поколения - величина нестабильная. Как это сказывается на расчетах в ДНК-генеалогии?
Действительно, в расчетах ДНК-генеалогии обычно оперируют поколениями, потому что при расчетах на один год константы скоростей мутации имели бы много нулей, а много нулей увеличивает вероятность ошибки. Например, для 12-маркерных гаплотипов (в стандартной системе FTDNA) величина константы скорости мутации равна 0.02 на гаплотип за условное поколение, или 0.0008 на гаплотип за год, или 0.0000666 на маркер за условное поколение. Условное поколение в контексте ДНК-генеалогии - это событие, которое происходит четыре раза в столетие. Численно и по времени оно близко к продолжительности поколения в житейском смысле этого слова, но не обязательно равно ему. Хотя бы потому, что продолжительность «бытового» поколения не может быть точно или хотя бы в среднем определена, она «плавает» в реальной жизни в зависимости от многих факторов, включая культурные, религиозные и бытовые традиции, примерно и в среднем от 18 до 36 лет, хотя границы этого не определены.
Такие организации, как ООН и ее подчиненные организации, как ЮНЕСКО и другие, как и прочие международные организации обычно принимают в своих подсчетах и рекомендациях продолжительность поколения в 25 лет, это же обычно принимают и демографы. Это все, разумеется, условности, и приняты только для некоторой стандартизации. Популяционные генетики рекоментациями ООН не руководствуются, и, в общем, правильно, но при этом используют продолжительности поколения кто во что горазд. Никакой научной основы в это не закладывают, поэтому уйдя от одних рекомендаций они никаких других не выработали. Недавно вышла статья по геномному анализу, там в расчетах применяли 29 лет на поколение, на каком основании - не пояснили. Другие берут от 18 до 35 лет на поколение.
В ДНК-генеалогии произвольные значения продолжительности поколений не используются, поскольку использовать столь «плавающую» величину для расчётов в широких временных диапазонах и для разных народов не представляется возможным или разумным. Исходя из этого положения, скорости мутаций откалиброваны под условно взятое поколение продолжительностью 25 лет. Если кому-то больше нравится 30 лет на поколение или любое другое количество лет, скорости можно перекалибровать, и в итоге окажутся ровно те же величины в годах. Так что сколько лет приходится на поколение - в данном случае не имеет значения, потому что при расчётах меньшему числу лет на поколение будет просто соответствовать пропорционально большее число поколений, и итоговая величина в годах не изменится.
Например, для 67-маркерных гаплотипов средняя константа скорости мутаций равна 0.12 на гаплотип на условное поколение в 25 лет. Поэтому при 120 мутаций на 50 гаплотипов общий предок этих 50 гаплотипов жил 120/50/0.12 = 20 условных поколений назад, то есть 20х25 = 500 лет назад. Если кому-то не нравится величина 25 лет на условное поколение (правда, причины такой нелюбви к 25 годам на поколение будут загадочными), и этот некто захочет использовать 30 лет на поколение, то нет проблем, просто константу скорости мутации придется пропорционально пересчитать, и она получится 0.12х30/25 = 0.144 мутаций на поколение в 30 лет. Тогда для 120 мутаций имеем 120/50/0.144 = 16.666 поколений по 30 лет, или те же 500 лет до общего предка. От чего ушли, к тому же и пришли.
Какие требования предъявляются к выборкам гаплотипов для расчетов в ДНК-генеалогии?
Как уже пояснялось, если взять любую выборку, тупо считать количество мутаций в гаплотипах, и разделить его на число гаплотипов и на константу скорости мутации, то что-то, конечно, получится. Именно так и проводят «расчеты» популяционные генетики. Но на самом деле там, скорее всего, получится некое бессмысленное число в поколениях или годах до якобы общего предка. Но откуда известно, что для данной выборки общий предок был один? А ведь так всегда и работают попгенетики, усредняя любые выборки. Поскольку после некоторых проб и ошибок ими было найдено, что таким образом происходит «умоложение» общих предков выборок, то попгенетики не нашли ничего лучшего, как втрое занизить величину константы скорости мутаций. Собственно, они ее «константой» никогда и не называли, просто «популяционная скорость мутаций». Вот так и работали - за счет усреднения время до общего предка занижали в неизвестное число раз, а за счет искусственного занижения «скорости мутаций» его опять увеличивали, правда, опять в неизвестное число раз, по принципу - «авось скомпенсируется». А оно не компенсировалось, а либо получалось значительно заниженным, либо значительно завышенным. Когда общий предок выборки был случайно один, то завышение времени до него составляло обычно 280-350%. Так и работали, и вот такими завышенными датировками заполонили академические журналы.
Честь нанесения такого ущерба науке, если попгенетику называть наукой, принадлежит Л.А. Животовскому, сотруднику Института общей генетики РАН. На основании какого-то вывиха здравого смысла, эту «популяционную скорость» узаконили после 2004 года, и более десяти лет в академических журналах гнали откровенный мусор. Широко известны «датировки» «общего предка» гаплогруппы R1a в Индии, которые Животовский с соавторами (эстонская школа попгенетики) загнали в 12-15 тысяч лет назад, когда они на самом деле составляют 3500-4000 лет назад, для разных выборок. Это на долгие годы не только затормозило изучение истории ариев в Индии, но и направило изучение истории по ложному пути.
Возвращаясь к исходному вопросу, только те мутации в гаплотипах имеет смысл рассчитывать, экстраполируя ко времени общего предка, которые подчиняются определённым количественным закономерностям. ДНК-генеалогия оперирует тремя экспериментальными факторами: 1) наличием снипов, относящих человека к определённому роду; 2) наличием мутаций, позволяющих оценивать время, прошедшее от общего предка совокупности гаплотипов и - при больших выборках - от начала самого рода, от самого далёкого из предков ныне живущих потомков данного рода (то есть здесь считаются сами мутации); 3) закономерностями переходов гаплотипов в их мутированные формы, без численного учёта самих мутаций (то есть здесь мутации не считаются, считаются немутированные гаплотипы).
Это позволяет оценить, насколько достоверны расчёты времени жизни общего предка по мутациям, и даёт ещё один, независимый способ расчетов. Методы, в которых считаются мутации, называются линейным, квадратичным и пермутационным, из которых наиболее прост линейный метод. Метод, в котором считаются немутированные гаплотипы, называется логарифмическим. В принципе, он использует формулы химической кинетики первого порядка (дополнительно вводя поправку на возвратные мутации). В линейном методе общее число мутаций в серии гаплотипов делится на число гаплотипов и на константу скорости мутации для данных гаплотипов. В логарифмическом методе берется логарифм отношения общего числа гаплотипов к числу немутированных гаплотипов, и делится на константу скорости мутации. Очень важно понять, что если линейный и логарифмический методы дают разные времена до общих предков, то выборка «кривая», то есть неоднородная, и происходит от нескольких общих предков, как минимум от двух. Получаемые времена до общих предков тогда лишены определенного смысла, предок «фантомный».
Приведем пример. Имеем неоднородную выборку из 300 гаплотипов в 25-маркерном формате, в которой 200 гаплотипов происходят от общего предка, который жил 3000 лет назад, и 100 гаплотипов - от предка, который жил 800 лет назад. Если мы построим дерево гаплотипов, то сразу увидим, что 200 гаплотипов образуют одну большую и глубокую (высокую) ветвь, и 100 гаплотипов - плоскую ветвь. Тогда сразу можно разделить ветви, и каждую из двух ветвей обсчитывать отдельно. В первой ветви из 200 гаплотипов все 25-маркерные гаплотипы будут разными (то есть логарифмический метод с ними работать не будет), что уже покажет, что общий предок жил более 2500 лет назад (при 2500 годах в выборке могли бы быть два одинаковых гаплотипа, потому что [ln(200/2)]/0.046 = 100 условных поколений, или 2500 лет до общего предка). На все 200 гаплотипов будет 1104 мутаций от базового гаплотипа, что даст 1104/200/0.046 = 120 условных поколений, или 120х25 = 3000 лет до общего предка (вкладом возвратных мутаций здесь для простоты пренебрегаем).
Во второй ветви из 100 гаплотипов в 25-маркерном формате 23 гаплотипа будут идентичными друг другу. Это даст [ln(100/23)]/0.046 = 32 условных поколений, то есть 800 лет до общего предка. Иначе говоря, если смешать эти 200 и 100 гаплотипов, то в этой смеси 23 гаплотипа будут одинаковыми (из молодой ветви), и мы получим [ln(300/23)]/0.046 = 56 условных поколений, то есть якобы 1400 лет до общего предка якобы одного для всех 300 гаплотипов. А мутаций во всей составной, неоднородной выборке будет 1251, если базовые гаплотипы обеих ветвей случайно совпадут. Тогда линейный метод даст 1251/300/0.046 = 91 условное поколение, или 2275 лет до якобы общего (фантомного) общего предка. Если базовые гаплотипы обеих ветвей будут разными, то время до фантомного общего предка будет практически любым между 3000 и 800 лет.
Итак, подводим итоги. Выборка составная, неоднородная, общие предки жили 3000 и 800 лет назад. Линейный метод дал фантомного общего предка 2275 лет назад, логарифмический - 1400 лет назад. То, что эти времена не совпали друг с другом, говорит о том, что так считать нельзя, и выборки надо разделять. Обычно разделение проводится с использованием деревьев гаплотипов.
Выборки в ДНК-генеалогии обычно несоизмеримо малые по сравнению с реальными величинами самих популяций. Как таким выборкам можно доверять? Есть «мнения», что выборки должны иметь как минимум 10 тысяч гаплотипов, чтобы получать относительно разумные данные.
Это совершенно ненаучная постановка вопроса, опять «по понятиям», по «мнениям», ни на чем не основанным. На это можно задать встречный вопрос - чтобы понять, что море соленое, нужно ли пробовать воду из всех бухточек? А как же химики делают анализ растворов, беря пробу всего лишь в долю миллилитра из ведра, цистерны, озера? Когда делают анализ крови, что, всю кровь из вас выкачивают? Нет. Значит, вопрос о размере пробы поставлен некорректно. Вопрос на самом деле сводится не к каким-то с потолка взятым числам, а к тому, равномерно ли перемешана система, чтобы по малой пробе, или выборке, судить обо всей системе. Этот вопрос в ДНК-генеалогии давно проработан, и результаты сопоставительных исследований показаны, например, в следующей таблице. Перед вами история определений времени до общего предка гаплогруппы R1a на «пост-советском пространстве» по разным сериям гаплотипов:
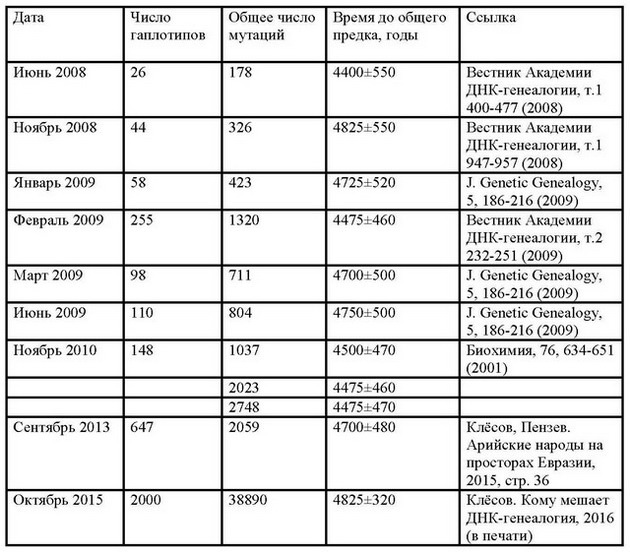
Из таблицы видно, что при последовательном переходе от серии из 26 гаплотипов до 2000 гаплотипов, и при увеличении числа мутаций в них в двести с лишним раз расчетное время до общего предка этих гаплотипов, в данном случае проживавшего на Русской равнине около пяти тысяч лет назад, получается практически одинаковым, в пределах погрешности расчетов. Это значит, что гаплотипы гаплогруппы R1a на пост-советском пространстве перемешаны достаточно хорошо, и все выборки вполне репрезентативны.
Конечно, могут быть и другие ситуации. Например, в горах в каждой долине состав гаплотипов (и гаплогрупп/субкладов) может заметно различаться, в отличие от того, что может наблюдаться на равнине, где гаплотипы более перемешаны за тысячелетия. Но в любом случае расчеты и выводы по гаплотипам относятся только к той конкретной выборке, которая рассматривалась, и только сопоставление разных выборок может показать, насколько система «перемешана», и решить вопрос об обобщении результатов расчетов и выводов на всю популяцию региона, или только ее конкретную часть. Но пока репрезентативность разных выборок наблюдается и в горах. Например, разные выборки осетин и карачаево-балкарцев дают воспроизводимые результаты для каждого из этих этносов. Перемешивание между ними обнаружено, но очень незначительное, практически не влияющее на результаты расчетов.
В большинстве случаев результаты расчетов почти не зависят от размера выборки (при числе гаплотипов больше двух-трех десятков), то есть они устойчивы к статистическим вариациям, если популяция достаточно перемешана и выбрка произведена неупорядочено. Размер выборки увеличивает точность, и то только до определенных пределов. Это, повторяем, относится к довольно большим популяциям, которые перемешались за тысячелетия, но именно с такими обычно и работают.
Что такое «деревья гаплотипов» в ДНК-генеалогии?
По ходу изложения в книге показываются деревья гаплотипов, которые отражают ДНК-картину этносов, родов, племен. Это вовсе не значит, что гаплотипы определяют этнос. Но они его в определенной степени описывают, каждый этнос по-разному. Человеку непривычному эти деревья представляются некими абстрактными образованиями, но в них на самом деле заключен большой смысл и в их графическом виде дается большой объем информации. Для того чтобы сделать деревья в этой книге более понятными и осмысленными, приведем несколько вводных, «модельных» примеров.
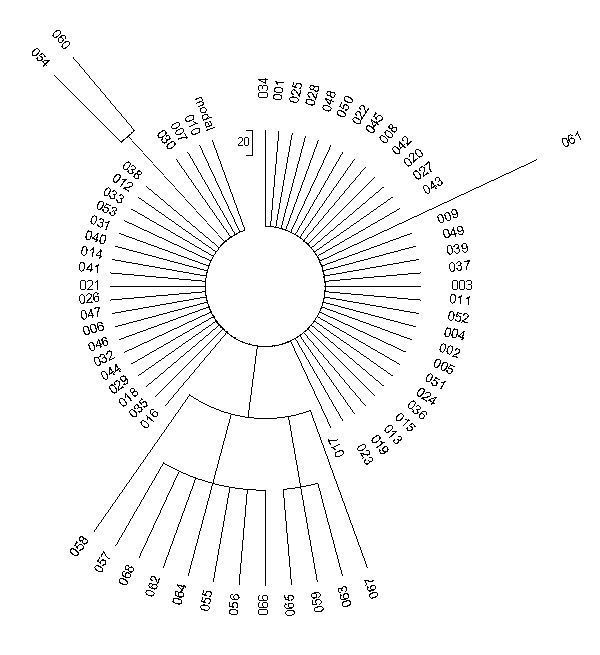
На рисунке приведено дерево реальной выборки из 68 гаплотипов гаплогруппы R1a. Гаплотипы простенькие, 6-маркерные, общий предок всех жил всего 625 лет назад, так что мутаций в гаплотипах набежало мало. Можно даже заранее посчитать, сколько. Это дерево гаплотипов потомков Джона, Лорда Британских островов, который жил в 14-м веке и умер в 1386 году. Его потомки - известный шотландский клан Мак-Доналдов (один из Мак-Доналдов был маршалом у Наполеона). Константа скорости мутации для 6-маркерных гаплотипов равна 0.0074 мутаций на гаплотип на условное поколение в 25 лет. Тогда за 625 лет (25 условных поколений) в каждом гаплотипе набежит 0.0074х25 = 0.185 мутаций, и на все 68 гаплотипов будет 0.185х68 = 13±4 мутаций. Погрешность рассчитывается по обычным правилам математической статистики.
Смотрим на дерево гаплотипов. Там - 15 мутаций, то есть в пределах погрешности. Можно посчитать самим, это все «вылезающие» из колеса спицы. А вылезают они в разных (и заранее непредсказуемых) местах, потому что мутации происходят неупорядоченно. 53 гаплотипа не мутированы, они образуют ровную гребенку, или «колесо» вокруг центральной части. Это - тот же самый гаплотип, что имел общий предок всех 68 человек, то есть Джон, Лорд Островов: 13 25 15 11 12 11.
53 человека этот гаплотип полностью сохранили, потому что 625 лет - это относительно малое время, и 15 мутаций - это все отклонения от предкового гаплотипа, что за это время смогли произойти. Показанные выше шесть чисел соответствуют шести участкам в Y-хромосоме ДНК, в которых повторяются выбранные исследователями короткие нуклеотидные последовательности. В первом участке - 13 повторов, во втором - 25 повторов, в третьем - 15 повторов и т.д. Мутация - это изменение числа повторов. Ошибся копирующий фермент при биологическом копировании Y-хромосомы, скопировал третий участок не 15 раз, как завещали предки, а 16 раз, получилось 13 25 16 11 12 11. Это - два идентичных гаплотипа, на дереве под номерами 054 и 060. С правой стороны - гаплотип 061, он имеет вид 14 25 15 11 12 11. У него мутация проскочила в первом участке, было 13 повторов, стало 14. То есть опять система копирования ошиблась в сторону завышения числа повторов. А вот в семерке идентичных гаплотипов под номерами 055-057, 062, 064, 066, 068 - мутация прошла во втором участке, на понижение (25 → 24) и получилось 13 24 15 11 12 11.
Действительно, этот участок, или маркер, как их называют, один из самых подверженных мутациям. Теория подсказывает, что в первом участке на все гаплотипы произойдет только одна мутация, во втором - семь мутаций, в третьем - две мутации. Так и получилось на практике. То есть даже при такой малой статистике закономерности в целом выполняются. Более того, все остальные пять мутаций на дереве гаплотипов относятся к тому же (второму) быстрому маркеру. Гаплотипы под номерами 059, 063 и 065 имеют вид 13 26 15 11 12 11, то есть мутация прошла на единицу на повышение, 25 → 26. Последние две мутации, в гаплотипах под номерами 058 и 067, оказались двойными (25 → 23, 25 → 27):
13 23 15 11 12 11
13 27 15 11 12 11
Они и сидят по обе стороны «букета», потому что не происходят одна из другой. Заметьте, что длина «спицы» в двух последних случаях тоже двойная, по сравнению со всеми остальными. То есть по виду дерева гаплотипов можно сказать, какие мутации одиночные, какие - двойные или тройные.
Внешний вид «дерева» определяется настройками профессиональной компьютерной программы, которая строит его на основании вводимого списка гаплотипов. Она сортирует гаплотипы по ветвям, то есть по похожести гаплотипов, сортирует по тому, в каких маркерах произошли мутации, сколько мутаций произошло, и по тому, насколько ветви древние. Поэтому по виду дерева человек опытный сразу может понять, какова структура той или иной популяции, сколько в ней основных ветвей, каков относительный возраст ветвей, и далее по числу мутаций в ветвях рассчитать, когда жил общий предок каждой ветви. Таких компьютерных программ много, наиболее известная - PHYLIP, с преобразованием MEGA.
По виду базовых гаплотипов в разных регионах и по возрасту их ветвей можно устанавливать, откуда, куда и когда проходили древние миграции людей и где сейчас живут их потомки. То есть можно проводить ДНК-генеалогическое картирование регионов, материков, и всей планеты, как в пространстве, так и во времени. В этом - методология ДНК-генеалогии. А дальше идут интерпретации получаемых данных в терминах истории, языкознания, антропологии, стыковка их с известными данными (тогда это по сути «калибровка»), или с данными неустоявшимися, сомнительными, конфликтными - тогда это дополнительная «точка опоры», или, наконец, это введение в научный оборот совершенно новых данных, и открытие пути для их верификации, проверки, обсуждения, выдвижения новых гипотез и положений.
Анатолий А. Клёсов, доктор химических наук, профессор