Кому мешает ДНК-генеалогия: Сопоставление гаплотипов гаплогруппы в Средней Азии и в Европе

Начало...
О своей новой книге «Кому мешает ДНК-генеалогия» профессор А.А. Клёсов, пишет, что её жанр оказался необычным. Это и учебник ДНК-генеалогии, и серия иллюстраций о достижениях ДНК-генеалогии, о ее открытиях и находках за последние годы, и срывание масок с лжецов и провокаторов, которым ДНК-генеалогия откровенно мешает. А в чем мешает - будет рассказано и показано. Пока же продолжаем вопросы и ответы по ДНК-генеалогии, которые занимают немалую часть новой книги.
Как современные гаплотипы могут указывать, когда и в какую сторону шли миграции тысячелетия назад?
Когда шли миграции (или пребывание данной популяции на рассматриваемой территории) - рассчитывается по временному расстоянию до общего предка популяции на данной территории, причем такие расчеты проводятся для той же гаплогруппы или субклада на разных территориях. Другими словами, проводится картирование территорий на максимально возможных расстояниях, по всему континенту или на нескольких континентах. Если вдоль пути миграции, то есть шлейфа миграций, идет систематическое уменьшение времен до общего предка, то миграция шла в сторону уменьшения времени. Пример - сопоставление серий гаплотипов гаплогруппы R1b в Средней Азии и в Европе показало, что общий их предок жил 16 тысяч лет назад. Но в Европе общий предок гаплотипов гаплогруппы R1b жил 4800 лет назад (на Пиренейском полуострове) и 4500 лет назад (в центральной Европе). Следовательно, миграции эрбинов, носителей гаплогруппы R1b, шли со стороны Средней Азии на запад, в сторону Европы.
Последующие исследования показали, что гаплогруппа R1b в районе Урала и на Русской равнине имела общих предков как минимум 7000 лет назад, на Кавказе - 6000 лет назад, в Месопотамии и на Ближнем Востоке - 5500 лет назад, на Пиренеях - 4800 лет назад. В ходе этого исследования направления миграций и их времена стали более понятны. Датировки ископаемых гаплотипов позволили подтвердить выводы, полученные при изучении современных гаплотипов - на территории ямной культуры (в Самарской области) были найдены костные останки носителей гаплогруппы R1b с археологическими датировками 5300 лет назад, а в Южной Сибири - костные останки носителя гаплогруппы R с датировкой 24 тысячи лет назад. В Европе (в Германии) были найдены ископаемые гаплотипы на территории культуры колокололовидных кубков, принадлежащие гаплогруппе R1b с датировкой 4500-4100 лет назад. Интересен и их состав по субкладам - M269, P310/L151, L51, P312, U152. Первый - обычный далекий родительский субклад из Сибири или ближнего Зауралья, два следующих - ближневосточные (образовались примерно 6200 и 5700 лет назад), P312 - ранний иберийский субклад, начальные колоколовидные кубки, U152 - его потомок (время образования примерно 4500 лет назад). Как видим, времена и направления миграции эрбинов, полученные поначалу с использованием гаплотипов современников, подтвердились на ДНК ископаемых костных останков.
Еще пример - датировки (по мутациям в гаплотипах) гаплогруппы R1a-Z93 уменьшаются от 5500 лет назад в Европе до 4050 лет назад в Индии (для субклада R1a-Z93-L657, которого в Европе еще не было), то есть по ходу миграционного маршрута появляются и новые, нисходящие снипы. И таких примеров можно приводить множество.
Оперирует ли ДНК-генеалогия понятием «раса»?
Понятия «раса» в ДНК-генеалогии нет. ДНК-генеалогические линии поднимаются из глубин тысячелетий, десятков тысяч лет, сотен тысяч лет, а расовые признаки отдельных людей могут кардинально меняться за несколько поколений. Дело в том, что расовые признаки являются продуктом отца и матери, и если родители принадлежат разным расам, признаки расплываются, и через несколько поколений могут измениться до неузнаваемости. Вот пример - как женитьба на русских женщинах всего в трех поколениях сделала из сибирских монголоидов (хакасов) внучек-европеоидов.



Еще пример - раскопки алтайской пазырыкской культуры позволили выснить, как скифы гаплогруппы R1a стали монголоидными всего за несколько поколений. Причина обратная той, что на фотографиях выше. Раскопки показали, что у этих скифов типичные монголоидные митохондриальные ДНК, то есть они женились на местных монголоидных женщинах, и через несколько поколений дети становились монголоидами, при наличии унаследованной по мужской линии гаплогруппы R1a. Иначе говоря, мужские гаплогруппы не определяют расу, и напротив, раса не определяет гаплогруппу. В этой ситуации понятие расы в ДНК-генеалогии ничего не дает.
Что могло вызывать массовые миграции в древности?
В самом общем виде ответ такой - поиски лучшей доли. Конкретные причины могли быть самыми разнообразными - это и передвижения за уходящим зверем, который давал пищу и одежду, и уход от превосходящих сил неприятеля, но самый мощный, видимо, фактор был резкое ухудшение климатических условий - суровые зимы, наводнения, засухи, надвигающиеся оледенения. В середине III тыс. до н.э., в результате заселения Европы эрбинами, носителями гаплогруппы R1b, разрушилась Старая Европа, погибли процветающие тысячелетиями балканские культуры, и в Европе древние гаплогруппы либо погибли, либо бежали на периферии континента. Это - гаплогруппы G2a, I2a, E1b-V13, R1a. Гаплогруппа R1a передвинулась на восток, на Русскую равнину, но только малым составом. Об этом говорит то, что в Европе древних гаплогрупп R1a практически не осталось, за исключением малых долей процента, а в Восточной Европе гаплогруппа R1a практически обнулилась, то есть прошла бутылочное горлышко популяции, и общий предок практически всех носителей R1a жил примерно 4600-4900 лет назад (по разным выборкам).
Произошли ли люди от обезьян?
Произошли, но не от тех обезьян, которых мы видим в зоопарке. Те, что в зоопарке, или резвятся на полянах в Африке или в Азии, или ходят по краю крыши домов в Индии - это продукты эволюционного развития от общих предков их и человека миллионы лет назад. Например, общий предок шимпанзе и человека жил более 4 миллионов лет назад, и тот, что сейчас в зоопарке - прошел свой эволюционный путь, видимо - не самый удачный. Мы, современные люди, произошли от того же общего предка, несколько более удачным образом, правда, не все. И не только с шимпанзе у нас был общий предок. Был общий предок и с гориллой, и с орангутангом, и с макакой. Вот как совпадают их нуклеотидные последовательности в ДНК.
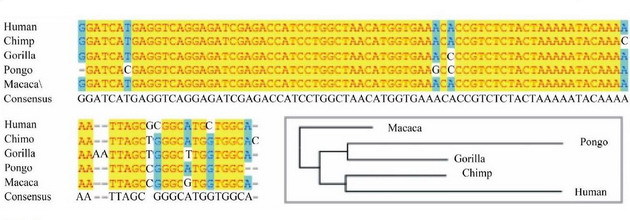
На рисунке - сопоставление нуклеотидных последовательностей фрагмента Y-хромосомы для современных человека, шимпанзе, гориллы, орангутанга и макаки. Как видно, из 97 нуклеотидов показанной последовательности Y-хромосомы у современного человека и шимпанзе не совпадают только четыре, то есть совпадение наблюдается в 96% нуклеотидов. Совпадение с гориллой - в 93% нуклеотидов. Случайных совпадений до такой степени быть не может, мы - родственники, правда, весьма отдаленные.
Многие, даже в научной литературе, педалируют тот факт, что мы не можем найти «пропущенные звенья» эволюции, и что это якобы ставит под сомнение теорию эволюции. Да, найти их было бы интересно, но приведенные выше данные по геному делают эти «пропущенные звенья» не такими уже и обязательными. Ясно, что они где-то лежат глубоко в земле, но всю землю пока не перекопали, и перекопают не скоро. Тем не менее, новые находки не прекращаются, и каждый раз непредсказуемые. Появятся и пропущенные звенья.
Коррелирует ли Y-хромосома с общим геномом?
Когда как. Часто коррелирует, иногда нет. Описано немало случаев, когда у двух родных братьев определяли гаплотипы Y-хромосомы, и они были, разумеется, одинаковы или практически одинаковы, а данные по аутосоме (то есть по рекомбинируемым хромосомам генома) заметно различались. И братья далеко не всегда были похожи друг на друга. Порой напротив, геномные данные были близки, а гаплотипы отличались друг от друга на тысячи лет (при переводе мутаций в хронологические показатели). Осложняет картину и то, что геномные данные разные исследователи представляют по-разному. И действительно, секвенирование геномов дает списки из десятков, сотен тысяч или миллионы снипов, и существует множество способов, как эти списки наглядно или не очень наглядно представлять. Каждый раз - это по сути поиск вариантов приближения или упрощения общей картины, подгонка приближения к ответу на заданный вопрос.
Есть ли «русская ДНК» или «русский геном»?
Говоря об отдельных людях - нет. Если говорить о большой группе этнических русских - в целом есть. И понятно, почему есть. Свои обычно, статистически, женятся на своих, как и выходят замуж. Вряд ли в русских деревнях встречали много чернокожих, монголов, австралийских аборигенов или американских индейцев. Поэтому в целом одни и те же огромные наборы снип-мутаций крутятся в популяции веками, вносимых мужчинами и женщинами этой популяции. Если построить карту мира, состоящую из таких популяций, то африканские будут совершенно отличны от монгольских, китайских, латиноамериканских или европейских. Здесь я пишу «европейских» более осторожно, потому что разойдутся ли они по разным «цветам», или останутся однородными по всей Европе зависит от того, как представлять данные, на каком уровне обработки и приближения. Нередко бывает, что, например, русских от французов не отличить, но если перейти на другой уровень обработки данных, то различаются.
Каковы перспективы геномного анализа популяций и их истории?
В целом перспективы у популяционного анализа геномных данных - но это все равно пока не ДНК-генеалогия - огромные, но пока мы находится в самом начале этого пути, идя методом проб и ошибок. Примитивизм обработки данных и получаемых выводов порой просто шокирует, хотя сами исходные данные определенно замечательны. Срезание углов и шапкозакидательство в науке никогда не помогали. К сожалению, у популяционных генетиков не принято при анализе генома давать погрешности расчетов, приводить альтернативные варианты интерпретаций данных и воздерживаться от мнений, которые в избытке вбрасываются в статьи. В итоге достоверность полученных интерпретаций часто не просто нулевая, а имеет отрицательную величину. Целый ряд рассмотрений подобных статей с «геномным анализом» приведен в моей авторской колонке на Переформате.
Например, недавно по миру прошла широковещательная информация, что геномные данные показали, что носители гаплогруппы R1b из ямной археологической культы (нынешняя Самарская область) принесли в Европу индоевропейские языки. Там что ни слово, то недоразумение. Во-первых, авторы не получили никаких данных, что носители ямной культуры (примерно 5300 лет назад), гаплогруппа R1b, говорили на индоевропейских языках, это было просто придумано. Да и как могли получить? Выкопаннные кости они и есть кости, они не говорят. Всё, что определили, это то, что ДНК в них относилась к гаплогруппе R1b. Далее, то, что носители той ДНК/R1b пришли из ямной культуры в Европу (и принесли ИЕ языки), было провозглашено только на том основании, что в Европе много гаплогруппы R1b. Иначе говоря, авторы просто провели прямую линию между ямной культурой и Европой. Они даже не посмотрели на то, что они нашли не просто R1b, а ее субклад R1b-Z2103/Z2105, которого в Европе почти нет. Миграционный путь субклада Z2103/Z215 был не в Европу, а на юг, в Месопотамию и на Ближний Восток, где этого субклада множество, причем с той же датировкой 5000-6000 лет назад. Но в Месопотамии сейчас нет индоевропейских языков, как нет нигде на миграционном пути гаплогруппы R1b из Южной Сибири в Европу, с основным входом через Пиренейский полуостров, где у басков имеется около 90% гаплогруппы R1b, и язык опять неиндоевропейский. Это, к сожалению, типичный пример, как популяционные генетики используют геномный анализ для «изучения истории».
Насколько точны (или неточны) расчеты в ДНК-генеалогии?
Все расчеты в ДНК-генеалогии производятся с погрешностями. Точные цифры получены быть не могут в принципе. Дело в том, что мутации неупорядоченные, поэтому мы имеем дело со статистикой. Чем больше выборка, тем точнее расчеты. Чем длиннее гаплотипы - тем точнее расчеты. Погрешность зависит от числа гаплотипов в выборке, от протяженности гаплотипов, от того, насколько точно определена и выверена, откалибрована константа скорости мутации, от того, насколько древний общий предок. Особенно неточно сравнивать мутации в парах гаплотипов, особенно гаплотипов коротких. На двух гаплотипах мутация-другая могла добавиться буквально в предыдущем или нынешнем поколении. Это может сразу привести к прибавлению-отнятию тысячи лет «в одном поколении». А в большой выборке разница относительно нивелируется статистикой. Есть выборки в тысячи гаплотипов - там расчеты, конечно, точнее. В ДНК-генеалогии часто важна не абсолютная точность, а концептуальный вывод.
Например, если некто утверждает, что носители гаплогруппы R1b (которые сейчас составляют примерно 60% мужского населения Западной и Центральной Европы), жили там, в Европе, еще 30 тыс. лет назад (а такими утверждениями популяционных генетиков, без расчетов, была заполнена академическая литература по данным вопросам до недавнего времени), а расчеты ДНК-генеалогии показали 4800 лет назад, то здесь дело не в точности, а в принципиальном выводе. То же самое по ДНК-генеалогии Кавказа - если данные показывают, что Кавказ заселялся носителями гаплогруппы J2 из Месопотамии примерно 7-6 тыс. лет назад, причем заселялся разными племенами и по разным территориям, и уже известно какими именно, и в какое время, то это дает важные данные историкам и археологам, которые продолжают горячо спорить по данным вопросам. То же самое по ДНК-генеалогии Прибалтики - данные показывают, что Прибалтика со стороны Финляндии и со стороны Южной Балтики заселялась всего 2000-2500 лет назад (со стороны Южной Балтики примерно на 500 лет раньше), но сами мигранты, которые на исходе миграции разделились на две большие ветви - северная и южная - имеют общего предка примерно 3575 лет назад, который, видимо, жил на Урале.
В какой степени гаплотипы определяются территориями и регионами?
Гаплотипы в немалой степени (но не всегда) связаны с определёнными территориями. Причина в том, что в древности большинство людей передвигались племенами, родами. Род, по определению, это группа людей, связанных родством, то есть происходят (по мужской линии) от одного общего предка. То есть гаплотипы у них обычно одинаковые или близкие. Проходили тысячелетия, численность родов порой сокращалась до минимума («бутылочное горлышко» популяции), и если выживший имел некоторую мутацию в гаплотипе, то его потомки уже «стартовали» с этой новой мутацией, копируя ее поколениями в своих ДНК, а мужчины - в своей Y-хромосоме. В популяционной генетике это называется «эффект основателя», что, в общем, особой дополнительной смысловой нагрузки не несет, это просто калька с английского языка, в котором это тоже не несет определенной информации. Некоторые люди покидали род по своей или чужой воле - плен, бегство, путешествия, военные походы, и выжившие начинали новый род на новом месте. В итоге карта мира с точки зрения ДНК-генеалогии получилась пятнистой, и каждое пятно порой имеет свой превалирующий гаплотип - гаплотип рода. Часто он и есть гаплотип предка, начавшего род на данной территории.
Получается, людей можно классифицировать по виду гаплотипов и снипов?
Да, это именно так. Например, носители гаплогрупп А и В сейчас в основном живут в Африке (но где эти гаплогруппы появились, пока непонятно, похоже, что не в Африке); гаплогруппа С встречается особенно часто среди монголоидов и жителей Австралии и Океании, хотя жители Австралии и Океании далеко не монголоиды (что показывает, в совокупности с другими данными, что первые носители гаплогруппы С были не монголоидны), подгруппы (субклады) далеко отклонились друг от друга в ходе развития или эволюции; гаплогруппу G находят в древней Европе (в основном ископаемые гаплотипы), на Кавказе, в Передней Азии, в Афганистане; гаплогруппа J - образовалась или в древнейшие времене перешла на Ближний Восток, и оттуда многие носители гаплогруппы J и ее субкладов J1 и J2 перешли Кавказ не позднее 6-7 тысяч лет назад; представителей гаплогруппы J1 довольно много на Кавказе, а также среди арабов и евреев на Ближнем Востоке, гаплогруппы J2 много на Кавказе, в Месопотамии и среди жителей Средиземноморья; гаплогруппа L появилась, похоже, в Передней Азии или на Ближнем Востоке, и разошлась в разные стороны - ее подгруппа (субклад) L1a - в Индии, L1b - в Грузии, L1c - в Афганистане, в северном Пакистане, на восточном Кавказе.
Гаплогруппу N имеют многие жители Китая, Сибири, севера России, Прибалтики и части Скандинавии, куда и прибыли носители гаплогруппы N с востока - Урала и до того Южной Сибири тысячелетия назад. Гаплогруппа I - возможно, имеет историю на Русской равнине продолжительностью не менее 40-45 тысяч лет, и после почти полного вымирания или истребления примерно 4500 лет назад сейчас почти исключительно находится в Европе, где возродилась относительно недавно, 3700-2300 лет назад (для I1 и части I2, соответственно).
Гаплогруппы R1a и R1b появились в Южной Сибири после долгой миграции их предков, носителей гаплогруппы К, из которой образовалась гаплогруппа Р и затем Q и R, возможно, из Европы или с Русской равнины на восток, далеко за Урал. Оттуда гаплогруппы R1a и R1b пришли в Европу, причем разными миграционными путями. R1a стала основной гаплогруппой Восточной Европы, R1b - западной и центральной Европы.
Данное положение сводится к тому, что у каждого мужчины есть снип из определённого набора, по которому можно отнести носителя к определённому древнему роду. Времена появления снипов, обычно применяемых в ДНК-генеалогии, относятся к диапазону от 25-40 тысяч лет для «старых» снипов до 10-15 тысяч лет для «молодых», но сейчас уже выявляют снипы, образовавшиеся всего 600-1000-1500 лет назад. Четверть всего ирландского мужского населения имеют снип (R1b-M222), который образовался примерно 4300 лет назад, но прошел бутылочное горлышко популяции в начале нашей эры, и общий предок этой четверти населения жил всего 1800 лет назад. В континентальной Европе снипа М222 практически нет.
Продолжение следует...
Анатолий А. Клёсов, доктор химических наук, профессор