А. Клёсов - Кто такие скифы, кто их предки и потомки? - 2
Окончание. Начало здесь.
Как видно, на первых 12 маркерах индийский брамин, действительно, отличается от восточных славян только парой 13-30 → 14-32
Оказалось, что эта пара, 14-32, характерна для многих гаплотипов субклада R1a-L342.2-L657, то есть более позднему субкладу в динамике мутаций юго-восточной ветви гаплогруппы R1a. Эта пара характерна для ариев Индии, Ирана, Ближнего Востока (ОАЭ, Бахрейн, Саудовская Аравия), то есть куда арии и доходили; расчетные датировки общих предков - те же 3500-4000 лет. Ниже - примеры современных гаплотипов их прямых потомков:
.
.
13 25 15 11 11 14 12 12 10 14 11 32 - Индия
13 25 15 10 11 14 12 13 10 14 11 32 - Иран
13 25 16 11 11 13 12 12 11 14 11 32 - ОАЭ
13 25 15 10 11 14 12 12 10 14 11 32 - Араб (страна не указана)
13 25 15 10 11 14 12 12 10 14 11 32 - Араб (страна не указана)
13 25 15 11 11 14 12 12 10 14 11 32 - Бахрейн
13 24 15 10 11 14 12 12 10 14 11 32 - Саудовская Аравия
.
13 25 16 11 11 14 Х Х 10 14 11 32 - Ископаемый гаплотип скифов, 3800-3400 лет
.
А у киргизов этот гаплотип является предковым для всей киргизской популяции гаплогруппы R1a-L342.2:
.
13 25 16 11 11 14 12 12 10 14 11 32 - 15 9 11 11 11 23 14 21 31 12 15 15 16
.
с общим предком, который жил 2100±250 лет назад. «Классические» времена скифов, конец прошлой эры. Получается, что киргизы гаплогруппы R1a (которой у них много) - прямые потомки древних скифов.
.
Вот и приходим к выводу, что в отношении происхождения родов и племен, гаплогрупп и субкладов в ДНК-генеалогии, понятия ариев, скифов, восточных славян в ряде контекстов взаимосвязаны и взаимозаменяемы. Мы просто относим их к разным временным периодам, и порой к разным территориям. Это именно мы относим, для упрощения рассмотрения, а скорее, на основе устоявшихся традиций исторической науки. Ясно, что киргизы - не славяне, как не славяне и арабы. Но все они - потомки арийских общих предков. Это ветви одного дерева. К этому вопросу мы вернемся в конце статьи. Поэтому ответ на вопрос - являются ли славяне потомками скифов? - будет таким. В ряде случаев - да, являются прямыми потомками; во многих случаях славяне и скифы - потомки одних и тех же общих предков, ариев, носителей гаплогруппы R1a.
.
Но ведь известно по археологическим данным, что среди скифов были монголоиды? Известно. Однако если бы у тех монголоидов были определены гаплогруппы, то с хорошей вероятностью у них тоже была бы гаплогруппа R1a. Как такое может быть? И вот здесь - новый виток сведений об алтайских ариях-скифах. Переходим к Пазырыкской археологической культуре и современным жителям Алтая с гаплогруппой R1a.
.
Причины монголоидности восточных скифов. Пазырыкская культура - это археологическая культура железного века (III-V вв. до н.э., хотя некоторые опускают дату до VI века до н.э.), которую относят к «восточному скифскому кругу». Регион - горный Алтай и смежные территории Алтая, Казахстана и Монголии. Основное занятие - кочевое скотоводство. Высказывается предположение, что пазырыкская культура - производная от афанасьевской.
.
Недавно изучались гаплотипы и гаплогруппы (мужские и женские) современных жителей этого региона (Dulik et al, 2012), и митохондриальные гаплогруппы (женские по сути, поскольку мужчины получают их от матери, но дальше не передают, нет митохондрий в сперматозоидах) ископаемых костных остатков пазырыкской культуры (Gonzalez-Ruiz et al, 2012). Оказалось, что большинство мужских гаплогрупп в регионе относятся к гаплогруппе R1a, причем наибольшее их количество было в народности алтай-кижи. R1a содержали также тубалары, челканы, кумандины. На втором месте по численности была гаплогруппа Q, затем С, потом N, остальные - минорные, единичные гаплогруппы, включая R1b, которые, как правило, случайны, и могли попасть туда в любое время.
.
Однако у носителей гаплогруппы R1a на Алтае была характерная особенность. Если на Русской равнине и в Центральной Европе у них преимущественно митохондриальная гаплогруппа (мтДНК) Н, как и у их жен и подруг, так называемая «европейская», или «западная» мтДНК, то на Алтае у R1a, в основном, восточные, «восточно-евразийские», азиатские мтДНК - A, C, D и G, их у носителей гаплогруппы R1a до половины и двух третей, остальные - минорные, единичные. Европеоидных мтДНК у них почти нет.
.
Носители мтДНК A, C, D и G, как правило - монголоидные женщины и их сыновья и дочери, опять же монголоидные. Вот и разгадка, почему алтайские носители гаплогруппы R1a, потомки ариев-скифов, как правило, сами монголоидны. Антропологию в значительной степени определяют женщины. К тому же, у алтайских R1a те же монголоидные женщины, видимо, сменили язык всей популяции на тюркский.
Анализ гаплотипов алтайцев с точки зрения ДНК-генеалогии проведен в статье (Клёсов, 2012).
.
Дерево гаплотипов имеет необычный вид:
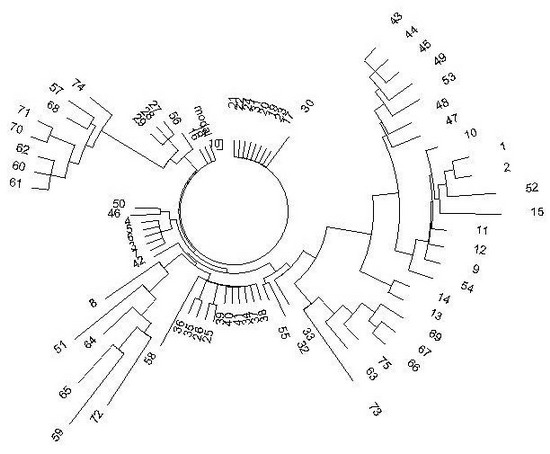
Дерево из 75 гаплотипов гаплогруппы R1a на Алтае в 17-маркерном формате. Верхняя левая ветвь - тубалары (57, 68, 70, 71, 74), челканы (60, 61, 62) и алтай-кижи (57). Нижняя правая ветвь - подобный состав: тубалары (66, 67, 69, 73, 75) и один челкан (63). Нижняя левая ветвь - смешанная: кумандины (64, 65), тубалар (72), алтай-кижи (8, 51, 59). Остальные гаплотипы - алтай-кижи.
.
Вид необычный потому, что гаплогруппа одна, то есть род один, а ветви расходятся в разные стороны, будучи заметно изолированными друг от друга. Это означает, что у данной популяции была тяжелая судьба. Племена погибали, немногие спасшиеся бежали, начинали свои генеалогические линии практически с нуля. Это повторялось, и опять спасались, бежали, и опять начинали свои линии. То есть повторялся эффект «последнего из могикан». В итоге все ветви на дереве гаплотипов относительно молодые, это набор относительно молодых «кустов», пересаженных со старых черенков, которые погибли. Но исходный общий предок жил в далекой древности, на это указывают масштабные расхождения между ветвями.
.
Для сравнения, дерево гаплотипов группы R1a на Русской равнине выглядят несравненно более благополучно:

Дерево из 257 гаплотипов гаплогруппы R1a на Русской равнине (по 12 областям Российской Федерации). Резко выделяющиеся гаплотипы Ar32 и Ar38 попали в список автора работы [Roewer, 2008] (и на построенное мной дерево) ошибочно, они относятся к другим гаплогруппам. Я не стал их снимать, чтобы показать, насколько дерево чувствительно к «чужакам» (Клёсов, 2009).
.
Еще более благополучно выглядит дерево гаплотипов Русской равнины для большого числа (801) протяженных гаплотипов (67-маркерных):

Дерево из 801 гаплотипа гаплогруппы R1a на Русской равнине - в 67-маркерном формате. Из статьи (Rozhanskii and Klyosov, 2012).
.
Посмотрим более внимательно на современные алтайские гаплотипы. Левая верхняя ветвь на рис. 3 из восьми гаплотипов (тубалары и челканы) имеет следующий предковый гаплотип:
.
13 24 16 9 12 14 10 14 11 32 14 14 20 12 17 11 23 (алтайские гаплотипы, ветвь)
.
В ветви всего 10 мутаций на 8 гаплотипов, то есть на 8х17 = 136 маркеров, что дает 10/8/0.034 = 37 → 38 условных поколений, то есть 950±315 лет до общего предка (стрелка - поправка на возвратные мутации, 0.034 - константа скорости мутации для 17-маркерных гаплотипов). Иначе говоря, общий предок этой ветви жил примерно в XI веке нашей эры, плюс-минус три века. Ясно, что он удален от скифов по времени, но гаплотип показывает, что современные алтайцы - прямые предки скифов. Тип гаплотипа тот же, та же самая пара 14-32.
.
13 25 16 11 11 14 10 14 11 32 15 14 20 12 16 11 23 (ископаемые скифы, R1a)
.
У подножия описанной ветви находится мини-ветвь из четырех гаплотипов популяции алтай-кижи, три из которых одинаковы, а четвертый (56) отличается всего на одну мутацию:
13 26 16 10 11 14 10 14 11 32 15 14 21 12 16 11 23 (№ 27, 28, 29)
13 25 16 10 11 14 10 14 11 32 15 14 21 12 16 11 23 (№ 56)
.
Мы видим, что у них та же самая характерная «подпись» - пара 14-32. Более того, у них всего три и две мутации, соответственно, от ископаемого скифского гаплотипа, то есть разницы почти нет.
.
Одна мутация между приведенными выше гаплотипами помещает их мини-ветвь на 1/0.034 = 29 → 30 поколений, то есть примерно на 750 лет назад. Но между этой мини-ветвью (алтай-кижи) и ветвью тубаларов и челканов - 6.5 мутаций, или 5900 лет между их общими предками. Это помещает их общего предка на (5900+950+750)/2 = 3800 лет назад. Это как раз датировка ископаемых гаплотипов скифов.
.
Иначе говоря, от ДНК-линии, идущей от 3800 лет назад (которая, в свою очередь, тоже явно прошла бутылочное горлышко популяции) остались две подветви с возрастом 950 и 750 лет назад. Но расстояние между ними выдает, что они далеко разошлись от их общего предка, и на сколько они разошлись друг от друга можно без труда вычислить. Это расстояние между общими предками двух алтайских ветвей и составляет указанные выше 5900 лет.
.
Гаплотипы, подобные древним скифским, имеют и остальные ветви дерева гаплотипов на рис. 1. Например, малая ветвь слева из семи гаплотипов (в которых всего три мутации):
.
13 25 16 11 11 14 10 14 11 32 - 15 14 21 10 16 11 23 (325 лет до общего предка)
.
Довольно древняя ветвь из 6 гаплотипов (на 7 часов) [27 мутаций]:
13 25 15 10 11 14/15 10 13 11 30/31 - 15 14 20 12 16 11 23 (3800 лет до общего предка)
.
Это вполне может быть субклад L342.2, базовый гаплотип которого на западе Русской равнины следующий:
13 25 16 11 11 14 10 13 11 30 - 15 14 20 12 16 11 23
.
Молодая ветвь из 10 гаплотипов внизу дерева, в которой всего 4 мутации:
13 25 16 11 11 14 10 14 11 32 - 15 14 21 12 17 11 23 (300 лет до общего предка)
.
Ветвь из семи гаплотипов в верхней правой части дерева, в которой всего 5 мутаций, что дает 5/7/0.034 = 21 поколение, или примерно 525 лет до общего предка:
13 26 16 10 11 17 11 14 11 32 - 15 14 19 11 15 11 23
.
Подветвь из 9 гаплотипов на 3 часа (в которой всего 7 мутаций, то есть общий предок жил 7/9/0.034 = 23 поколения, то есть 575 лет назад), с базовым гаплотипом:
13 26 16 11 11 17 11 14 11 31 - 15 14 19 11 15 11 23
.
Видно, что это родственная ветвь предыдущей. У них одинаковые значения большинства аллелей, и они отличаются всего двумя мутациями, то есть их общие предки расходятся на 2/0.034 = 59 → 63 поколения, то есть на 1575 лет. Их общий предок жил (1575+525+575)/2 = 1340 лет назад. Ясно, что эта двойная ветвь - молодая (в отношении своего общего предка). Видно, как фрагментированы ветви, как они рассыпаются на совсем недавно выжившие и давшие недавних же потомков.
.
Основной вывод - что эти современные алтайские гаплотипы, точнее, их носители - потомки древних скифов, они же древние арии, с предком на Русской равнине.
.
Раскопки пазырыкской культуры выявили три мтДНК, относящиеся к бронзовому веку и шестнадцать мтДНК - к железному веку. К сожалению, Y-хромосомные ДНК не изучались, но мы уже знаем, что они с большой вероятностью дали бы, в основном, гаплогруппу R1a. Но и полученная информация оказалась важной. Из всех 19 ископаемых мтДНК гаплогрупп 11 оказались азиатскими (A, C, D, и G), и 8 - западными, точнее, западно-евразийскими (HV, J, U, T, K). Все три гаплогруппы бронзового века оказались азиатскими.
.
Гаплогруппы железного века дали смесь европейских и азиатских гаплогрупп. Это в любом случае показывает, что скифы были и европеоидные, и монголоидные, причем арии-скифы, пришедшие в алтайский регион в бронзовом веке, то есть самые ранние, брали в жены местных монголоидных женщин, и их потомки, сохраняя гаплогруппу R1a, были уже монголоидными. Это опять объясняет монголоидность некоторых (или многих) скифов, кочевавших по степям Евразии. Но многие скифы, очевидно, откочевывали на восток со своими европейскими женами и подругами, что и дало «западные» мтДНК в их потомках, в том числе и на Алтае в древности. Вот и получалось разнообразие антропологии скифов-кочевников, от европеоидных до монголоидных, при наличии основной гаплогруппы R1a.
.
Остальные известные сведения о скифах, а также мифы и легенды скифов и о скифах изложены во множестве источников, от античных до современных, мы на них поэтому останавливаться не будем. Мы уже знаем, что в отношении происхождения родов и племен, гаплогрупп и субкладов в ДНК-генеалогии, понятия ариев, скифов, восточных славян взаимосвязаны и взаимозаменяемы, мы просто относим их к разным временным периодам. И опять - это мы относим, для упрощения рассмотрения или на основе устоявшихся традиций исторической науки.
.
Скажем, при рассмотрении американских индейцев такого расслоения нет, они «нативные американцы» хоть 16 тысяч лет назад, хоть сейчас. И у древних скандинавов нет, они скандинавы тогда и скандинавы сейчас. И у древних германцев нет, они германцы в древности, и германцы (Germans) сейчас. А у населения Русской равнины, предков нынешних славян, историю разорвали разными названиями, да и по ним идут непрекращающиеся споры.
.
Критерии почему-то диктует лингвистика, хотя прекрасно известно, что при уходе в древность лингвистические критерии смещаются, ломаются, потому что языки к древности прогрессивно меняются, а потом вообще уходят как в песок, реконструируются только отдельные фрагменты, да и то весьма произвольно. Ну, как можно использовать критерии лингвистики при древности родов и племен более 4 тысяч лет назад, не говоря о 6 тысячах лет и более? Вот так и превратились арии в неких безликих «индоевропейцев», языки которых ныне разошлись по всему миру, и к древним ариям уже в большинстве случае не имеют никакого отношения.
.
На самом деле древние арии на Русской равнине и были славянами, если судить по пантеону (языческих) богов, которые перекликаются от Восточной Европы через Русскую равнину до Индостана, а также сказаниям и мифам. Их топонимы и гидронимы были древними славянскими просто по определению. И не нужно сравнивать их звучания с современными славянскими, язык с тех пор изменился, и не он должен лежать в основе классификации древних племен и народов.
.
А вот характерные «подписи» в их ДНК не изменились, и унаследованы без принципиальных изменений в ходе многих тысячелетий и десятков тысяч лет, только естественным путем разветвились, оставляя общую картину доступной для несложной реконструкции. Языки здесь - вторичный признак, не главный, они изменчивы и принципиально подвержены произвольным толкованиям и интерпретациям лингвистов. Которые, кстати, и сами между собой не могут договориться в подавляющем большинстве случаев.
.
И вот если мы по всей справедливости будем считать языки в данном контексте вторичными факторами, то картина вырисовывается вполне четкая: арии, скифы и восточные славяне - это одни и те же люди, в своей естественной хронологической динамике. Большинство их принадлежало и принадлежит одному и тому же роду - R1a.
.
Теперь вопрос - а насколько «большинство»? Полагаю, точная цифра здесь не имеет значения. Ясно, что в их составе были и другие гаплогруппы, но они не доминировали. Во всяком случае, данных таких нет, будут - рассмотрим. У гаплогруппы R1b была своя славная история, но в составе ариев, прибывших в Индию, их не было. Во всяком случае, в современной Индии их очень мало, в высших кастах почти нет, и среди 367 браминов, тестированных на гаплогруппы, не было найдено ни одного случая гаплогруппы R1b (Sharma et al, 2009). Мы не можем исключить наличие некоторого количества эрбинов, носителей гаплогруппы R1b, среди скифов, но что это даст? Ну, допустим, были…
.
Что дальше? Да и среди современных этнических русских носителей гаплогруппы R1b примерно 5%. Для сравнения, гаплогруппы R1a среди этнических русских - до двух третей в южных областях - Курской, Белгородской, Орловской. В среднем, по всем европейским областям Российской Федерации, включая северные (во многом финно-угорские) - половина R1a от всего населения.
.
Не было среди ариев и скифов и носителей гаплогруппы N. У тех была другая история, тоже славная, просто по определению. Они со стороны Южной Сибири ушли на север примерно 8 тысяч лет назад, затем повернули на запад, и через Урал, став по дефинициям лингвистов уграми, разошлись на несколько ветвей. Одна ветвь через Поволжье ушла в центральную Европу, стала венграми, хотя в Венгрии сейчас их осталось очень мало, единицы процентов.
.
Возможно, и в древности было столько же. Другая ушла на Балтику, разошлась на финскую (N1c1-Z1935), балтийскую (N1c1-L1022) и южно-балтийскую (N1c1-L550) ветви. Ни одна из них к ариям или скифам отношения не имеет, хотя из двух последних вышло много славян (по определениям лингвистов) гаплогруппы N1c1. В наше время их среди этнических русских примерно 14%, но доходит до половины на Русском севере. На юге России - единицы процентов южных балтов и финно-угров (по происхождению).
.
То же относится и к носителям гаплогруппы I (I1 и I2), не было их среди ариев или скифов. Почти все они были уничтожены в Центральной Европе в III тыс. до н.э. (между 4800 и 4000 лет назад), в ходе заселения эрбинами европейского континента. Остатки носителей гаплогруппы I бежали на Британские острова и на Карпаты, и начали возрождаться только 3600 лет назад (I1) и 2300 лет назад (I2). Для арийских миграций это было уже поздно, так что носители гаплогруппы I в Индию или Иран не попали, как не попали и на Ближний Восток (немного есть, но относительно недавние). Они остались в пределах Европы, в основном Атлантической ее части (I1 и I2), в Скандинавии (I1), и на Балканах (I2). Поэтому в составе скифов их не было, тем более что начало возрождения гаплогруппы I2 на Карпатах - это уже конец прошлой эры, времена излета скифов в том виде, как их представляет академическая история.
.
В принципе, в составе скифов могла быть представлена гаплогруппа Q, поскольку ее имеют сибирские и монгольские народы (хотя у последних - гаплогруппы Q всего 6%). Единственное основание к этому, кроме аргумента «по общим понятиям», это наличие относительно небольшого количества гаплогруппы Q в современной Европе, да и то на малом уровне: 2% в Венгрии, 2% в Румынии, 1% во Франции. Хотя это все могут быть потомки угров, пришедших в Венгрию уже в нашей эре, и разошедшихся по Европе. Исходя из довольно большого притока скифов в Европу, можно заключить, что среди них гаплогруппа Q была представлена совсем мало. В общем, так и получается, что скифами были в основном арии, носители гаплогруппы R1a. А то, что они были разные по антропологии, от европеоидных до монголоидных, мы уже объяснили на основании данных ДНК-генеалогии.
.
Еще соображение, вытекающее из сказанного выше. Все известные деления скифов на «царских скифов», «скифов-пахарей», «скифов-воинов», «скифов-кочевников», «скифов-земледельцев», «борисфенитов» (Геродот) и прочих являются поверхностными. По подобным «признакам» мы и сегодняшних россиян можем делить на «пахарей», «воинов», «инженеров», «профессоров», «медицинских работников» и прочих, но относится ли это деление к происхождению русского народа? Хотя делить и изучать можно и нужно, и этим занимаются социальные службы, но надо понимать, с какими целями, для чего это стоит делать, и на какие загадки истории - в случае скифов - это ответит.
.
Литература
Клёсов, А.А. (2009) Гаплотипы восточных славян: девять племен? Вестник Российской Академии ДНК-генеалогии, т. 2, № 2, 232-251.
Клёсов, А.А. (2012) Гаплотипы группы R1a на Алтае: «автохтоны» и «индоевропейцы». Вестник Академии ДНК-генеалогии, т. 5, № 12, 1511-1525.
Dulik, M.C., Zhadanov, S. I., Osipova, L.P., Askapuli, A., Gau, L., Gokcumen, O., Rubinstein, S., Schurr, T.G. (2012) Mitochondrial DNA and Y chromosome variation provides evidence for a recent common ancestry between Native Americans and indigenous Altaians. Amer. J. Human Genetics, 90, 1-18. DOI 10.1016/ajhg.2011.12.014.
Gonzalez-Ruiz, M., Santos, C., Jordana, X., Simon, M., Lalueza-Fox, C., Gigli, E., Aluja, M., Malgosa, A. (2012) Tracing the origin of the East-West population admixture in the Altai region (Central Asia). PLOS One, 7, 1-11. e48904.
Gray, R.D. and Atkinson, Q.D. (2003) Language-tree divergence times support the Anatolian theory of Indo-European origin. Nature, 426, 435-439.
Haak, W., Brandt, G., de Jong, H.N., Meyer, C., Ganslmeier, R., Heyd, V., Hawkesworth, C., Pike, A.W.G., Meller, H., Alt, K.W. (2008) Ancient DNA Strontium isotopes, and osteological analyses shed light on social and kinship organization of the Later Stone Age. Proc. Natl. Acad. Sci. U.S. 105, 18226-18231.
Keyser, C., Bouakaze, C., Crubezy, E., Nikolaev, V.G., Montagnon, D., Reis, T., Ludes, B. (2009) Ancient DNA provides new insight into the history of south Siberian Kurgan people. Human Genetics 126, 395-410.
Klyosov, A.A., Rozhanskii, I.L. (2012) Haplogroup R1a as the Proto Indo-Europeans and the legendary Aryans as witnessed by the DNA of their current descendants. Adv. Anthropol. 2, No. 1, 1-13.
Klyosov, A.A., Rozhanskii, I.L. (2012) Haplogroup R1a as the Proto Indo-Europeans and the legendary Aryans as witnessed by the DNA of their current descendants. Adv. Anthropol. 2, No. 2, 1-13.
Rozhanskii, I.L., Klyosov, A.A. (2012) Haplogroup R1a, its subclades and branches in Europe during the last 9000 years. Adv. Anthropol. 2, No. 3, 139-156.
Sharma, S., Rai, E., Sharma, P., Jena, M., Singh, S., Darvishi, K., Bhat, A.K. et al. (2009) The Indian origin of paternal haplogroup R1a1* substantiates the autochtonous origin of Brahmins and the caste system. J. Human Genet. 54, 47-55.
Анатолий А. Клёсов,
доктор химических наук, профессор
Источник