Африканские сказки - 2
Часть 2-я. Начало здесь.
В первой части я кратко рассмотрел археологические аспекты постулата "выхода из Африки" и аргумент "генетического разнообразия" на который опираются приверженцы этой мейнстримной, и ставшей почти догматической, концепции.
Но максимальное генетическое разнообразие не обязательно может совпадать с местом происхождения. Представим себе что неандертальцы (а заодно и денисовцы) не вымерли бы после минимального генетического обмена с хомо сапиенс, а широко перемешались бы с ним так что генетика человечества покрывала бы весь спектр этих видов. Это привело бы к разнообразию намного большему чем сегодня наблюдается в Африке, но к месту происхождения человека это не имело бы отношения. Так что разнообразие является не следствием региона происхождения, а того что много достаточно далеко отстающих друг от друга ветвей выжили в этом регионе параллельно друг с другом, без тотального доминирования одной узкой ветви. Подобное произошло в Африке, где современное генетическое разнообразие сложилось из многих волн миграции из Евразии, выживших параллельно в течении сотен тысяч лет.
В Евразии, после "бутылочного горлышка" 70 тлн, вероятно появились несколько новых мутаций, которые обеспечили их носителям значительное эволюционное преимущество, что привело к быстрому распространению и тотальному доминированию этой ветви на всем евразийском пространстве, и вымиранию или уничтожению всех других человеческих особей. Следствием этого явилось относительно низкое генетическое разнообразие евразийцев. Как и почему это произошло - является, наверное, одной из главных загадок в ранней истории человечества.
Рассмотрим некоторорые вопросы генетического разнообразия более подробно.
В значительной мере с точки зрения генетики постулат "выхода из Африки" основан на африканском происхождении субклада L3 митохондриальной ДНК. Это ключевой субклад который является предковым по отношению к гаплогруппам M и N, охватывающим почти все не-африканское человечество. L3 встречается как в Африке, так и в Евразии. Остальные субклады L имеют очень мало присутствия за пределами Африки, а гаплогруппы M и N - наоборот, имеют бесспорное евразийское происхождение и мигрировали в Африку позже.
Но... в последние два десятилетия появляется всё больше исследований, показывающих что L3 образовался за пределами Африки. Более того, даже не близко к Африке (на Ближнем Востоке или в Аравии), а очень далеко от неё - примерно в центре Азии или даже к востоку от него. Есть уже немало академических работ, выводы из которых должны быть сенсационными. В промежутке времени 70-60 тлн происходил достаточно мощный поток хомо сапиенс... в Африку, а не из Африки. Но "выход из Африки" именно в этом временном интервале - основополагающий постулат "африканского происхождения" человечества!
В большинстве этих статей в предисловиях дежурно упоминается про "хорошо установленный факт африканского происхождения" современного человека, но основные выводы в этих статьях прямо противоречат этому. Речь идёт не только о субкладе L3, но параллельно с ним исследования Y-хромосомных ДНК подтверждают миграцию в Африку в этом интервале, связанном с гаплогруппой Е (по современной классификации), образовавшейся примерно одновременно с L3 и вместе с этим субкладом мигрировавшей в Африку.
Ниже подобраны только несколько заголовков статей на эту тему:

В этих статьях предлагаются различные формы преодоления очевидного противоречия с "африканским происхождением современного человека", такие как сдвиг главного "выхода из Африки" на период 120-125 тлн вместо канонического 60-70 тлн, или общие рассуждения о двусторонней миграции туда-сюда, предполагающием плотные встречные потоки через один или два узких перешейка (Синай и Баб-эль-Мандеб). Ни то ни другое обьяснение не убедительно.
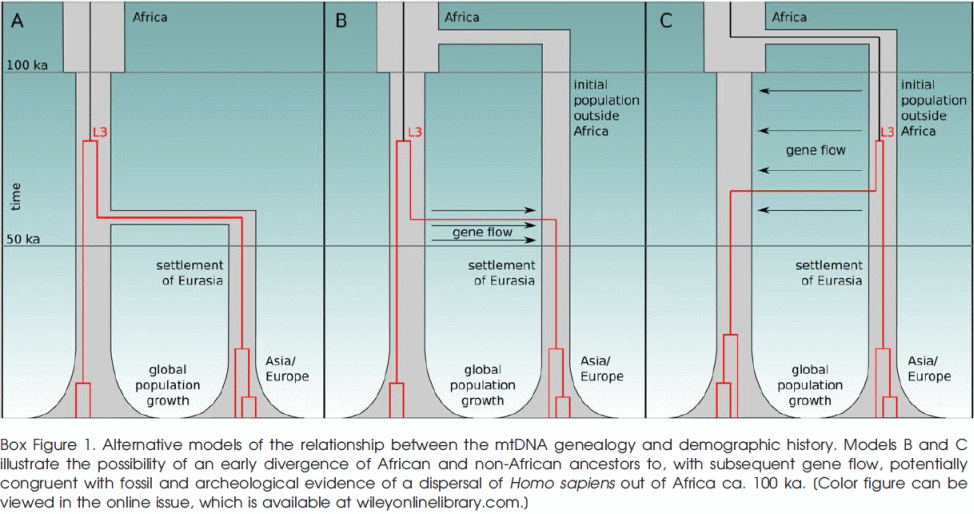
Наконец, в статье "Rethinking the Dispersal of Homo sapiens out of Africa", Evolutionary Anthropology 24:149-164 (2015) авторы, на основании большого массива данных по ветвям L3 с высоким разрешением, моделируют несколько сценариев представленных на картинке ниже:
A) образование L3 в Африке и единовременный "выход из Африки" около 60 тлн;
B) Первоначальный выход из Африки около 125 тлн, образование L3 в Африке около 70-80 тлн и второй выход из Африки около 60 тлн генетически смешавшийся с потомками первого выхода;
C) Выход из Африки около 125 тлн, образование L3 в Евразии и обратный поток в Африку 60-70 тлн.
Выяснилось что лучше всего удовлетворяет имеющимся данным сценарий C, хуже - A и гораздо хуже - B. Но сценарий A к сегодняшнему дню полностью несостоятелен, так как существование хомо сапиенс вне Африки намного раньше чем 60 тлн многократно доказано. Так что сценарий C выигрывает с большим преимуществом, подтверждая что субклад L3 образовался далеко от Африки и в интервале 60-70 тлн. происходила миграция в Африку, а не из неё. Осталось только убрать дежурный "первоначальный выход из Африки" кочующий из статьи в статью по политическим соображениям...
Похожую картину представляют и другие древние гаплогруппы. Если взять гаплогруппы C и D Y-хромосомы (предшествующие E и тем более поздним I, J, и другим), их наибольшее распространение сегодня наблюдается очень далеко от Африки: немногочисленные представители гаплогруппы D живут от Тибета до Бирмы, очень многочисленная C широко распространена в восточной и северной Азии, причем самые древние субклады её коррелируют с удалением от Африки.
Но вернемся к митохондриальной гаплогруппе L. Далее я предложу собственную интерпретацию, не взятую из академических источников, поэтому кто хочет может обвинять меня в дилетантстве.
Посмотрим на картy сегодняшнего распространения в Африке основных субкладов митохондриальной макро-гаплогруппы L (в основном взято из Википедии). Для каждой из них на этих картах указаны предковые группы, и примерное время возникновения этих субкладов. В разных источниках можно найти разные оценки этого времени, но в целом приведенные числа примерно соответствуют схеме отсюда, взятой с мейнстримного сайта Антропогенез.ру, из статьи С. Боринской, критикующей Клёсова (и в которой по-прежнему утверждается африканское происхождение L3).
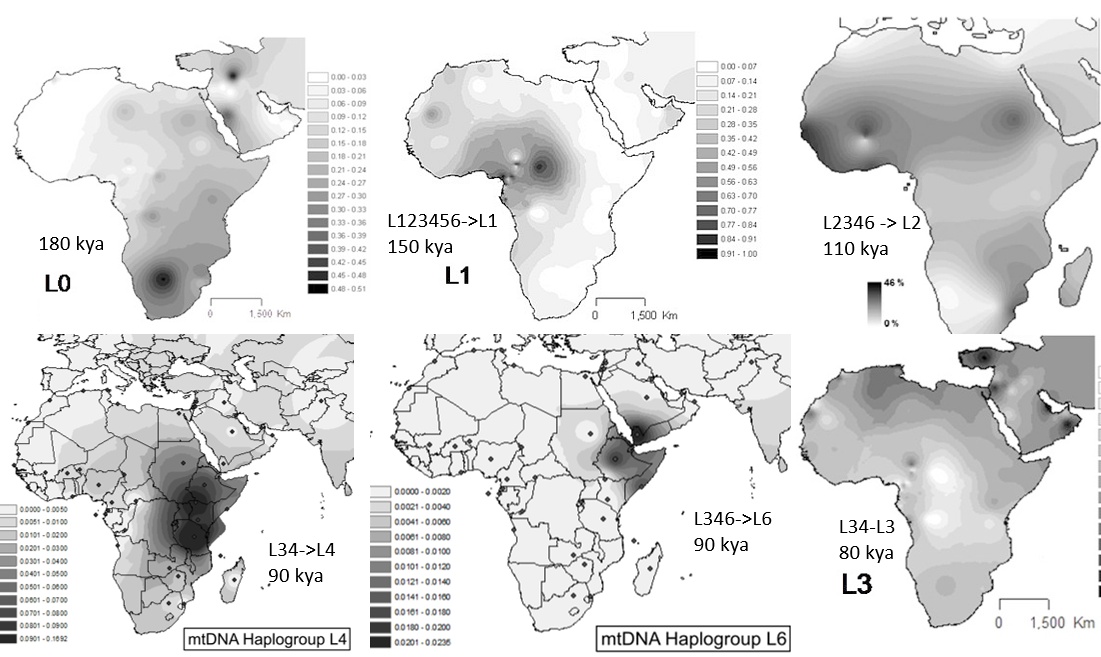
Картина распространения основных субкладов митохондриальной гаплогруппы L плохо соответствует модели возникновения их в Африке, но прекрасно сочетается с моделью отдельных, разделенных многими тысячелетиями, волн миграций, приходящих в Африку из Евразии через узкие перемычки - Синайский полуостров и Баб-эль-Мандебский пролив. Про некоторые из этих волн можно довольно определенно сказать через какой из двух перешейков они прошли в Африку, про другие - менее определенно, или, вероятно, переход осуществлялся через оба из них.
Демографическая эволюция в каждой из этих волн миграций имела важные общие черты. Каждая последующая волна осуществлялась всё более современными хомо сапиенсами, более конкурентоспособными в эволюционной борьбе. В результате мигранты предыдущих волн вытеснялись из более благоприятных регионов в менее привлекательные и приспособленные для жизни. Демографические следы предыдущих миграций практически полностью изчезали в районах близких к перешейкам (Синай и Баб-Эль-Мандеб), но могли сохраняться в менее привлекательных регионах далеко от них.
Приведенная выше карта хорошо обьясняется примерно следующим сценарием.
L0 Это самая древняя ветвь, отделившаяся от остального человечества около 160-200 тлн. Сегодня этот митохондриальный субклад имеет максимальное присутствие среди койсанских племен (бушменов, готтентотов), общающихся на щелкающих языках - наименее развитых южноафриканских народоностей. Переход группы носителей этого субклада произошел, по-видимому, через Баб-эль-Мандеб, хотя небольшая часть могла пересекать и Синай. Эта группа распространилась в восточной Африке и на всей большой территории южнее экватора. По-видимому, L0 никогда не присутствовали в западной Африке, куда их привела бы миграция вдоль южного берега Средиземного моря, так что группа, прошедшая через Синай, была малочисленной или её не было вообще. Последующие волны миграций вытеснили группу L0 в наименее привлекательные районы её ареала, с центром в пустыне Калахари, где они и обитают сегодня. Но и группа L0, в свою очередь, вытеснила или уничтожила предшествующих хомо, живших в южной части Африки.
L1 Эта группа пришла в Африку позже 150 тлн, возможно в период эемского потепления (и соответствующего демографического подьёма всего вида хомо сапиенс около 130-120 тлн). По-видимому они распространились по Африке достаточно широко, в основном в западной, северной и центральной её частях. Но последующими волнами они были вытеснены в намного более ограниченные регионы - в густые джунгли центрально-западной Африки (сегодняшние Камерун, Центрально-Африканская республика, Конго) и пустыни Чада и Нигера и, далее на запад,
Мали и Мавритании. Сегодня этот субклад наиболее распространен среди пигмеев африканских джунглей.
L2 Волна, принесшая субклад L2, скорее всего пришла через оба перешейка в период около 100-90 тлн, и распространилась шире по континенту чем предыдущие волны L0 и L1, по всей западной, северной и юго-восточной Африке. Эта волна вытеснила предыдущих мигрантов из большинства регионов, оставив при этом пустыню Калахари койсанским племенам L0 и экваториальные джунгли - пигмеям L1. Но и она сама впоследствии была оттеснена из многих относительно благоприятных регионов последующими миграциями - из районов, примыкающих к перешейкам, из восточно-африканской саванны, из умеренно-климатической южной оконечности, из южного берега Средиземного моря.
L4/L5 Группа субклада L4, и малочисленного близкого к ней L5, пришла в Африку на несколько тысяч лет позже L2, по-видимому, только через южный перешеек. Она закрепилась в основном в центрально-африканской саванне (Кения, Танзания, Уганда), где она вытеснила потомков миграции L2. В южную и центральную Африку её проникновение оказалось незначительным.
L6 Этот немногочисленный субклад отделился от предкового субклада L3'4'6' около 100 тлн, вероятно, на Аравийском полуострове или на Ближнем Востоке. Впоследствии он был вытеснен более поздними сублкадами (L3 и его потомками) в изолированные горные районы Йемена. Часть представителей этого субклада перебралась через Баб-эль-Мандеб и закрепилась в пустынных регионах Эфиопии и Сомали.
L3 Наконец, этот субклад который стал предковым всему евразийскому человечеству. Он широко развился после "бутылочного горлышка" около 70 тлн и стал проникать в Африку, по-видимому, через оба перешейка, но в большей степени через северный. Эта волна в основном вытеснила предыдущие миграции из наиболее привлекательных регионов - южного побережья Средиземного моря, побережья Атлантики, южной оконечности континента. Эта волна, по-видимому, уже достаточно широко смешивалась с последующими миграциями, принесшими в Африку почти все современные гаплогруппы, о чем свидетельствует диффузная картина её нынешнего распространения.
Посмотрим теперь на то как нынешнее распространение гаплогруппы L в Африке обьясняется в существующих моделях "выхода из Африки". Ниже на картинке показаны два варианта этой модели - с более традиционным "возникновением L3 в Африке" (слева) и более новой концепцией возникновения L3 в Азии и миграции её "обратно в Африку" (справа). Я нарочно не даю ссылок, подобные картинки с различными небольшими варианциями можно найти во многих оригинальных и обзорных работах.
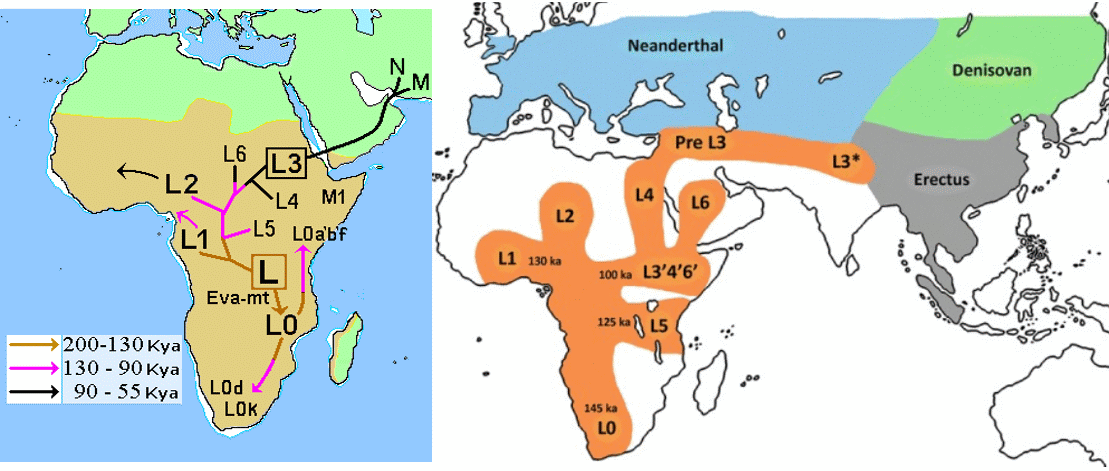
В первом случае появление "митохондриальной Евы" предполагается где-то в джунглях Мозамбика, и затем носители гаплогруппы L долго скитались по самым труднодоступным регионам Африки с низкими емкостями экологических ниш - густым джунглям и пустыням. В течении почти 100 тыс лет от этих миграций отделялись группы давшие, начало новым субкладам. Наконец, добравшись кружным путем до региона нынешней Эфиопии, как раз чтобы "прыгнуть" в Евразию, здесь появился субклад L3. Он тут же переправился через Баб-эль-Мандеб на Аравийский полуостров и стремительно распространился по всей Евразии (но не по Африке, где он был изначально), став предком всего не-африканского человечества. Приведенная схема миграций намного хуже обьясняет нынешнюю географию африканских субкладов L по сравнению с моделью последовательных волн из Азии через два перешейка, вытесняющих предыдущие волны в более отдаленные и менее привлекательные места.
Вторая схема, где L3 возникает далеко за пределами Африки, не менее натянута в плане географических перемещений. Она так же предполагает длительные скитания ранних субкладов L по самым суровым пустыням и джунглям, выход из Африки группы L3'4' через Синай, её стремительное продвижение до центра Азии, возникновение там L3 и такого же стремительного отката назад, в Африку, потомков L3. Она плохо сочетается с нынешней географией субклада L4, а так же L2.
Скорее всего я не первый кто заметил что распределение субкладов L хорошо согласуется с моделью отдельных "волн вытеснения" миграций из Евразии, а не непрерывного образования этих субкладов внутри Африки. Но подобная модель сегодня наверняка не будет принята в академической среде (мне проще, я не делаю научную карьеру в антропологии или генетике).
В следующей части (возможно, последней) я расмотрю свидетельство того что генетический обмен между хомо сапиенс и неандертальцами имел место очень давно - 350-400 тлн, что свидетельствует о присутствии ранних хомо сапиенс в это время вместе с неандертальцами, т.е. в Евразии, а не в Африке.
В первой части я кратко рассмотрел археологические аспекты постулата "выхода из Африки" и аргумент "генетического разнообразия" на который опираются приверженцы этой мейнстримной, и ставшей почти догматической, концепции.
Но максимальное генетическое разнообразие не обязательно может совпадать с местом происхождения. Представим себе что неандертальцы (а заодно и денисовцы) не вымерли бы после минимального генетического обмена с хомо сапиенс, а широко перемешались бы с ним так что генетика человечества покрывала бы весь спектр этих видов. Это привело бы к разнообразию намного большему чем сегодня наблюдается в Африке, но к месту происхождения человека это не имело бы отношения. Так что разнообразие является не следствием региона происхождения, а того что много достаточно далеко отстающих друг от друга ветвей выжили в этом регионе параллельно друг с другом, без тотального доминирования одной узкой ветви. Подобное произошло в Африке, где современное генетическое разнообразие сложилось из многих волн миграции из Евразии, выживших параллельно в течении сотен тысяч лет.
В Евразии, после "бутылочного горлышка" 70 тлн, вероятно появились несколько новых мутаций, которые обеспечили их носителям значительное эволюционное преимущество, что привело к быстрому распространению и тотальному доминированию этой ветви на всем евразийском пространстве, и вымиранию или уничтожению всех других человеческих особей. Следствием этого явилось относительно низкое генетическое разнообразие евразийцев. Как и почему это произошло - является, наверное, одной из главных загадок в ранней истории человечества.
Рассмотрим некоторорые вопросы генетического разнообразия более подробно.
В значительной мере с точки зрения генетики постулат "выхода из Африки" основан на африканском происхождении субклада L3 митохондриальной ДНК. Это ключевой субклад который является предковым по отношению к гаплогруппам M и N, охватывающим почти все не-африканское человечество. L3 встречается как в Африке, так и в Евразии. Остальные субклады L имеют очень мало присутствия за пределами Африки, а гаплогруппы M и N - наоборот, имеют бесспорное евразийское происхождение и мигрировали в Африку позже.
Но... в последние два десятилетия появляется всё больше исследований, показывающих что L3 образовался за пределами Африки. Более того, даже не близко к Африке (на Ближнем Востоке или в Аравии), а очень далеко от неё - примерно в центре Азии или даже к востоку от него. Есть уже немало академических работ, выводы из которых должны быть сенсационными. В промежутке времени 70-60 тлн происходил достаточно мощный поток хомо сапиенс... в Африку, а не из Африки. Но "выход из Африки" именно в этом временном интервале - основополагающий постулат "африканского происхождения" человечества!
В большинстве этих статей в предисловиях дежурно упоминается про "хорошо установленный факт африканского происхождения" современного человека, но основные выводы в этих статьях прямо противоречат этому. Речь идёт не только о субкладе L3, но параллельно с ним исследования Y-хромосомных ДНК подтверждают миграцию в Африку в этом интервале, связанном с гаплогруппой Е (по современной классификации), образовавшейся примерно одновременно с L3 и вместе с этим субкладом мигрировавшей в Африку.
Ниже подобраны только несколько заголовков статей на эту тему:
В этих статьях предлагаются различные формы преодоления очевидного противоречия с "африканским происхождением современного человека", такие как сдвиг главного "выхода из Африки" на период 120-125 тлн вместо канонического 60-70 тлн, или общие рассуждения о двусторонней миграции туда-сюда, предполагающием плотные встречные потоки через один или два узких перешейка (Синай и Баб-эль-Мандеб). Ни то ни другое обьяснение не убедительно.
Наконец, в статье "Rethinking the Dispersal of Homo sapiens out of Africa", Evolutionary Anthropology 24:149-164 (2015) авторы, на основании большого массива данных по ветвям L3 с высоким разрешением, моделируют несколько сценариев представленных на картинке ниже:
A) образование L3 в Африке и единовременный "выход из Африки" около 60 тлн;
B) Первоначальный выход из Африки около 125 тлн, образование L3 в Африке около 70-80 тлн и второй выход из Африки около 60 тлн генетически смешавшийся с потомками первого выхода;
C) Выход из Африки около 125 тлн, образование L3 в Евразии и обратный поток в Африку 60-70 тлн.
Выяснилось что лучше всего удовлетворяет имеющимся данным сценарий C, хуже - A и гораздо хуже - B. Но сценарий A к сегодняшнему дню полностью несостоятелен, так как существование хомо сапиенс вне Африки намного раньше чем 60 тлн многократно доказано. Так что сценарий C выигрывает с большим преимуществом, подтверждая что субклад L3 образовался далеко от Африки и в интервале 60-70 тлн. происходила миграция в Африку, а не из неё. Осталось только убрать дежурный "первоначальный выход из Африки" кочующий из статьи в статью по политическим соображениям...
Похожую картину представляют и другие древние гаплогруппы. Если взять гаплогруппы C и D Y-хромосомы (предшествующие E и тем более поздним I, J, и другим), их наибольшее распространение сегодня наблюдается очень далеко от Африки: немногочисленные представители гаплогруппы D живут от Тибета до Бирмы, очень многочисленная C широко распространена в восточной и северной Азии, причем самые древние субклады её коррелируют с удалением от Африки.
Но вернемся к митохондриальной гаплогруппе L. Далее я предложу собственную интерпретацию, не взятую из академических источников, поэтому кто хочет может обвинять меня в дилетантстве.
Посмотрим на картy сегодняшнего распространения в Африке основных субкладов митохондриальной макро-гаплогруппы L (в основном взято из Википедии). Для каждой из них на этих картах указаны предковые группы, и примерное время возникновения этих субкладов. В разных источниках можно найти разные оценки этого времени, но в целом приведенные числа примерно соответствуют схеме отсюда, взятой с мейнстримного сайта Антропогенез.ру, из статьи С. Боринской, критикующей Клёсова (и в которой по-прежнему утверждается африканское происхождение L3).
{kind=link}
Картина распространения основных субкладов митохондриальной гаплогруппы L плохо соответствует модели возникновения их в Африке, но прекрасно сочетается с моделью отдельных, разделенных многими тысячелетиями, волн миграций, приходящих в Африку из Евразии через узкие перемычки - Синайский полуостров и Баб-эль-Мандебский пролив. Про некоторые из этих волн можно довольно определенно сказать через какой из двух перешейков они прошли в Африку, про другие - менее определенно, или, вероятно, переход осуществлялся через оба из них.
Демографическая эволюция в каждой из этих волн миграций имела важные общие черты. Каждая последующая волна осуществлялась всё более современными хомо сапиенсами, более конкурентоспособными в эволюционной борьбе. В результате мигранты предыдущих волн вытеснялись из более благоприятных регионов в менее привлекательные и приспособленные для жизни. Демографические следы предыдущих миграций практически полностью изчезали в районах близких к перешейкам (Синай и Баб-Эль-Мандеб), но могли сохраняться в менее привлекательных регионах далеко от них.
Приведенная выше карта хорошо обьясняется примерно следующим сценарием.
L0 Это самая древняя ветвь, отделившаяся от остального человечества около 160-200 тлн. Сегодня этот митохондриальный субклад имеет максимальное присутствие среди койсанских племен (бушменов, готтентотов), общающихся на щелкающих языках - наименее развитых южноафриканских народоностей. Переход группы носителей этого субклада произошел, по-видимому, через Баб-эль-Мандеб, хотя небольшая часть могла пересекать и Синай. Эта группа распространилась в восточной Африке и на всей большой территории южнее экватора. По-видимому, L0 никогда не присутствовали в западной Африке, куда их привела бы миграция вдоль южного берега Средиземного моря, так что группа, прошедшая через Синай, была малочисленной или её не было вообще. Последующие волны миграций вытеснили группу L0 в наименее привлекательные районы её ареала, с центром в пустыне Калахари, где они и обитают сегодня. Но и группа L0, в свою очередь, вытеснила или уничтожила предшествующих хомо, живших в южной части Африки.
L1 Эта группа пришла в Африку позже 150 тлн, возможно в период эемского потепления (и соответствующего демографического подьёма всего вида хомо сапиенс около 130-120 тлн). По-видимому они распространились по Африке достаточно широко, в основном в западной, северной и центральной её частях. Но последующими волнами они были вытеснены в намного более ограниченные регионы - в густые джунгли центрально-западной Африки (сегодняшние Камерун, Центрально-Африканская республика, Конго) и пустыни Чада и Нигера и, далее на запад,
Мали и Мавритании. Сегодня этот субклад наиболее распространен среди пигмеев африканских джунглей.
L2 Волна, принесшая субклад L2, скорее всего пришла через оба перешейка в период около 100-90 тлн, и распространилась шире по континенту чем предыдущие волны L0 и L1, по всей западной, северной и юго-восточной Африке. Эта волна вытеснила предыдущих мигрантов из большинства регионов, оставив при этом пустыню Калахари койсанским племенам L0 и экваториальные джунгли - пигмеям L1. Но и она сама впоследствии была оттеснена из многих относительно благоприятных регионов последующими миграциями - из районов, примыкающих к перешейкам, из восточно-африканской саванны, из умеренно-климатической южной оконечности, из южного берега Средиземного моря.
L4/L5 Группа субклада L4, и малочисленного близкого к ней L5, пришла в Африку на несколько тысяч лет позже L2, по-видимому, только через южный перешеек. Она закрепилась в основном в центрально-африканской саванне (Кения, Танзания, Уганда), где она вытеснила потомков миграции L2. В южную и центральную Африку её проникновение оказалось незначительным.
L6 Этот немногочисленный субклад отделился от предкового субклада L3'4'6' около 100 тлн, вероятно, на Аравийском полуострове или на Ближнем Востоке. Впоследствии он был вытеснен более поздними сублкадами (L3 и его потомками) в изолированные горные районы Йемена. Часть представителей этого субклада перебралась через Баб-эль-Мандеб и закрепилась в пустынных регионах Эфиопии и Сомали.
L3 Наконец, этот субклад который стал предковым всему евразийскому человечеству. Он широко развился после "бутылочного горлышка" около 70 тлн и стал проникать в Африку, по-видимому, через оба перешейка, но в большей степени через северный. Эта волна в основном вытеснила предыдущие миграции из наиболее привлекательных регионов - южного побережья Средиземного моря, побережья Атлантики, южной оконечности континента. Эта волна, по-видимому, уже достаточно широко смешивалась с последующими миграциями, принесшими в Африку почти все современные гаплогруппы, о чем свидетельствует диффузная картина её нынешнего распространения.
Посмотрим теперь на то как нынешнее распространение гаплогруппы L в Африке обьясняется в существующих моделях "выхода из Африки". Ниже на картинке показаны два варианта этой модели - с более традиционным "возникновением L3 в Африке" (слева) и более новой концепцией возникновения L3 в Азии и миграции её "обратно в Африку" (справа). Я нарочно не даю ссылок, подобные картинки с различными небольшими варианциями можно найти во многих оригинальных и обзорных работах.
В первом случае появление "митохондриальной Евы" предполагается где-то в джунглях Мозамбика, и затем носители гаплогруппы L долго скитались по самым труднодоступным регионам Африки с низкими емкостями экологических ниш - густым джунглям и пустыням. В течении почти 100 тыс лет от этих миграций отделялись группы давшие, начало новым субкладам. Наконец, добравшись кружным путем до региона нынешней Эфиопии, как раз чтобы "прыгнуть" в Евразию, здесь появился субклад L3. Он тут же переправился через Баб-эль-Мандеб на Аравийский полуостров и стремительно распространился по всей Евразии (но не по Африке, где он был изначально), став предком всего не-африканского человечества. Приведенная схема миграций намного хуже обьясняет нынешнюю географию африканских субкладов L по сравнению с моделью последовательных волн из Азии через два перешейка, вытесняющих предыдущие волны в более отдаленные и менее привлекательные места.
Вторая схема, где L3 возникает далеко за пределами Африки, не менее натянута в плане географических перемещений. Она так же предполагает длительные скитания ранних субкладов L по самым суровым пустыням и джунглям, выход из Африки группы L3'4' через Синай, её стремительное продвижение до центра Азии, возникновение там L3 и такого же стремительного отката назад, в Африку, потомков L3. Она плохо сочетается с нынешней географией субклада L4, а так же L2.
Скорее всего я не первый кто заметил что распределение субкладов L хорошо согласуется с моделью отдельных "волн вытеснения" миграций из Евразии, а не непрерывного образования этих субкладов внутри Африки. Но подобная модель сегодня наверняка не будет принята в академической среде (мне проще, я не делаю научную карьеру в антропологии или генетике).
В следующей части (возможно, последней) я расмотрю свидетельство того что генетический обмен между хомо сапиенс и неандертальцами имел место очень давно - 350-400 тлн, что свидетельствует о присутствии ранних хомо сапиенс в это время вместе с неандертальцами, т.е. в Евразии, а не в Африке.