Индейцы, где ваши гены?
Ещё о генетике народов, а также Расология американцев
Еще раз о дене-кавказских языках: Северо-восточный след
Переселения народов / Статья 2014 года
Обсуждение гипотезы о языке «эрбин», относившимся к гипотетической же дене-кавказской макросемье, вызвало такую бурную дискуссию, что в ней стало уже сложно ориентироваться. Некоторые тезисы участников обсуждения повторяются раз за разом, и уже трудно уследить за основной мыслью. ©Ещё в цикле Народы, где ваши гены?

Большое каноэ тлингитов, репродукция с гравюры 18-го века
Постараюсь разорвать порочный круг и дополнить материал по дене-кавказской гипотезе анализом самой дальней ветви этих языков - на-дене. Точнее, их носителей, живущих на Аляске и северо-западе Канады.
С тех пор, как дене-кавказская гипотеза была впервые сформулирована С. Старостиным в 1984 году, было собрано довольно много языкового материала, подтверждающего, по мнению ее сторонников, родство языков на-дене, по меньшей мере, с енисейскими. Однако все эти аргументы повисают в воздухе, если нет информации, когда и каким образом эти языки попали в Америку. Датировки выделения ветви на-дене приходятся на времена, когда уже образовался Берингов пролив, а Америка была заселена носителями других америндских языков. Отсюда закономерно следует предположение, что предки нынешних атапасков, навахо и воспетых в вестернах апачей переселились из Азии в Америку по воде, несколько ранее или примерно в то же время, что и предки эскимосов.
Насколько мне известно, археологи не находят однозначных свидетельств такого перехода, и лингвистам приходится опираться только на данные этнографии по сходству материальной культуры народов Аляски и Сибири. Однако датировок из такого материала не извлечь, а потому независимых подтверждений о родстве языков на-дене с другими языками гипотетической макросемьи практически нет.
Популяционные генетики в своих исследования коренных народов Америки также не находят доказательств нескольких волн миграции, но их выводы были сделаны в отсутствие данных по народам на-дене, не говоря уж о, мягко говоря, спорных методах их расчетов. Недавно появилась возможность рассчитать датировки, используя данные по ДНК коренных народов Аляски и северо-западной Канады, опубликованные в статьях C. Davis с соавторами 2011 года и M. Dulik с соавторами 2012 года. Всего было опубликовано 233 Y-хромосомных гаплотипа различных племен на-дене в 17- и 19-маркерном форматах, а также 363 гаплотипа из трех этнических групп эскимосов: юпик, инувиалук и инупиат. Как это уже неоднократно наблюдалось для других групп аборигенов, многие из их представителей оказались потомками европейских поселенцев по своей прямой мужской линии. Это около половины индейцев на-дене и чуть меньше четверти эскимосов. Оставшиеся 120 «чистокровных» на-дене и 264 эскимоса распределились по гаплогруппам С и Q.
В статье Dulik с соавторами в популяции эскимосов был обнаружен новый снип гаплогруппы Q (NWT01), на основании которого авторы выделили новый субклад Q1a6 и рассчитали время жизни общего предка его носителей - около 7500 лет назад. Казалось бы, чем не датировка переселения в Америку в послеледниковую эпоху? Увы, считали с помощью «эволюционных скоростей», а потому датировка не имеет ничего общего с реальностью. Еще спустя какое-то время нашли, что новый субклад Q1a6 оказался в не слишком дальнем родстве с ветвью, имевшей тогда обозначение Q1a1 (М120). Вот как выглядит соответствующий фрагмент древа гаплогруппы Q после уточнения:

Напомню, что американские индейцы относятся к другой ветви этой гаплогруппы, а именно Q1a2a1 (L54), и их общий предок с найденной у эскимосов генеалогической линией уходит не менее чем на 20000 лет назад. Родство последней с дальневосточной ветвью М120 дает возможность предположить, что мы видим след миграции в Америку, прошедшей после того, как растаял ледник, и перешеек между Чукоткой и Аляской скрылся под водой. Чтобы проверить это предположение, надо рассчитать, как давно жил общий предок двух ветвей, во время существования Берингии или после.
Для начала рассмотрим древо 17-маркерных гаплотипов гаплогруппы Q из выборок на-дене, эскимосов и народов Дальнего Востока, представителей ветви М120.
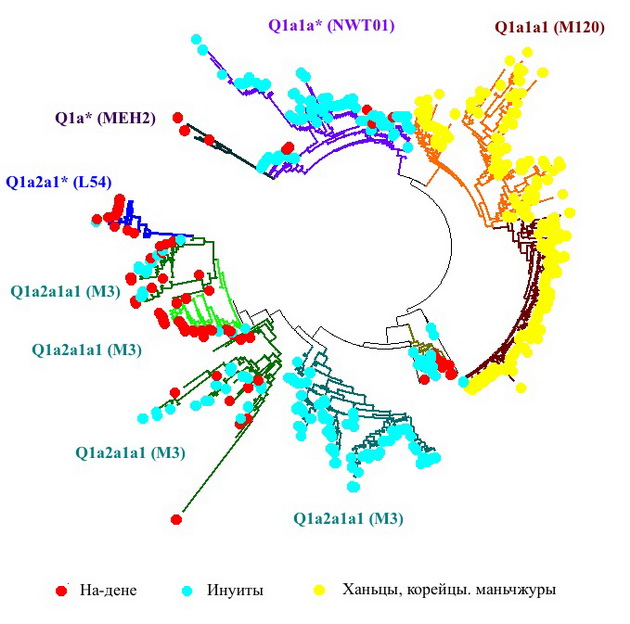
Аляскинская NWT01 и дальневосточная М120 располагаются на диаграмме ближе друг к другу, как и следовало бы ожидать. Первая из них сходится к предку, жившему 2500±300 лет назад, и имеет такой базовый 19-маркерный гаплотип в формате YFiler, с добавленными DYS426 и DYS388:
14 24 13 10 13 19 12 12 11 14 15 30 - 14 15 18 10 15 11 22
Вторая, М120, состоит из двух подветвей. Та, что выделена на диаграмме оранжевым цветом, имеет «возраст» 4000±450 лет и базовый гаплотип:
14 24 13 9 15 22 12 12 11 14 14 30 - 16 14 19 10 16 12 22
Ветвь, выделенная коричневым, соответственно, 3700±560 лет и
14 24 13 6 16 20 12 12 11 13 14 30 - 16 14 19 10 17 12 22
С учетом того, что очень редкое значение DYS391=6 (четвертый слева маркер) во второй ветви появилось, очевидно, в результате одной многошаговой мутации, эти две ветви расходятся на 7 мутаций, что соответствует дистанции в 6450 лет. С учетом «возраста» базовых гаплотипов это дает (4000 + 3700 + 6450)/2 ≈ 7100 лет до их общего предка. Сравним теперь эти базовые гаплотипы с ветвью, найденной у эскимосов Аляски. Получаем дистанции в 13 и 16 мутаций соответственно, что при пересчете дает общего предка, жившего около 11000 или 13500 лет назад, при оценке по первой и второй ветви. В пределах погрешности для столь давних времен это одно и то же. Для сравнения, европейские и американские линии субклада Q1a2a1 (L54) разошлись около 17000 лет назад, открытие прохода в ледовом щите между Аляской и территорией современного штата Монтана датируется временем около 13000 лет назад, а возникновение Берингова пролива - 11000 лет назад.
Было бы желательно уточнить датировку общего предка, используя протяженные гаплотипы, но пока таких данных нет. На сегодняшний момент можно сделать вывод, что предок генеалогических линий субклада Q1a1a из Китая и с Аляски жил, очевидно, в Азии тогда, когда Северную и (возможно) Южную Америку уже населяли предки современных индейцев, а сухопутный проход или ушел под воду, или значительно сократился по своей площади. Следовательно, ДНК-генеалогия дает весомые аргументы в пользу того, что предки современных эскимосов мигрировали из Азии в Америку позже, чем предки американских индейцев, скорее всего, водным путем. Когда именно это произошло, ни археология, ни ДНК-генеалогия, ни, тем более, лингвистика не дают однозначного ответа.
Верхний предел можно оценить, исходя из анализа других генеалогических линий эскимосов. В исследуемой выборке к ветви Q1a1a* (NWT01) относятся 115 из 264 эскимосских гаплотипов из аборигенных ветвей, то есть 44%. Еще 139 принадлежат к субкладу Q1a2a1a1 (M3), тому, что доминирует у индейцев как Северной, так и Южной Америки, но практически не встречается в Старом Свете. Очевидно, в этногенезе эскимосов приняли участие не только люди, пересекшие Берингов пролив в пост-ледниковую эпоху, но и потомки тех, кто осваивал север Америки несколькими тысячелетиями ранее. Из нескольких ветвей субклада М3, найденных в выборке, самая большая по численности (86 гаплотипов) состоит исключительно из эскимосов. На диаграмме она занимает ее нижнюю часть. Предок этой моноэтнической ветви жил 4000±460 лет назад, а ее базовый гаплотип выглядит следующим образом:
14 24 13 10 16 18 12 12 13 14 14 30 - 17 15 21 11 16 10 22
Поиск в доступных базах данных не выявил каких-либо близких к ней гаплотипов американских индейцев. Это, скорее всего, означает, что носители этой ветви влились в состав прото-эскимосов на самом раннем этапе этногенеза. Поскольку субклад М3 по всем признакам зародился в Америке, то это означает, что самое позднее 4000 лет назад предки современных эскимосов уже жили на севере Америки. Эту датировку можно рассматривать как подсказку для археологов в их интерпретациях древних культур Арктики.
Однако пока что нет ответа на поставленную задачу - доказать или опровергнуть факт миграции предков на-дене из Азии в послеледниковую эпоху. В ветви NWT01 народы на-дене представлены единичными гаплотипами, в основной же своей массе они принадлежат к различным ветвям субклада М3, что на диаграмме располагаются в левом нижнем секторе. Как уже говорилось выше, субклад М3 характеризует практически все группы американских индейцев, и в этой части своих генеалогических линий на-дене ничем не выделяются на их фоне. Если ограничиваться анализом гаплогруппы Q, то вопрос о второй волне миграции из Азии для предков на-дене можно считать закрытым - в их Y-ДНК ее следы отсутствуют. В свою очередь, этот факт ставит под серьезное сомнение их языковое родство с енисейскими, сино-тибетскими и т.д. Даже самые изощренные методы компаративистики не в силах распознать родство языков, разошедшихся 17000 лет назад или ранее.
А если искать следы общего происхождения за пределами гаплогруппы Q? На-дене выделяются на фоне других коренных народов Америки тем, что помимо гаплогруппы Q у них в заметной пропорции представлена гаплогруппа С. В данной выборке это ровно ⅓ (40 из 120-ти) индейцев из этнических групп атабасков, в числе которых племена тличо, кучин, тлингитов и т.д. Из этих 40 человек 37 относятся к ветви, что характеризуется снипом Р39. В текущей нотации она обозначается как C2b1, и на дереве гаплогруппы С располагается в том же субкладе C2b (L1373), что и ветви, широко представляющие у монголов, казахов, киргизов и других народов Центральной Азии. Вот как выглядит их общее дерево.
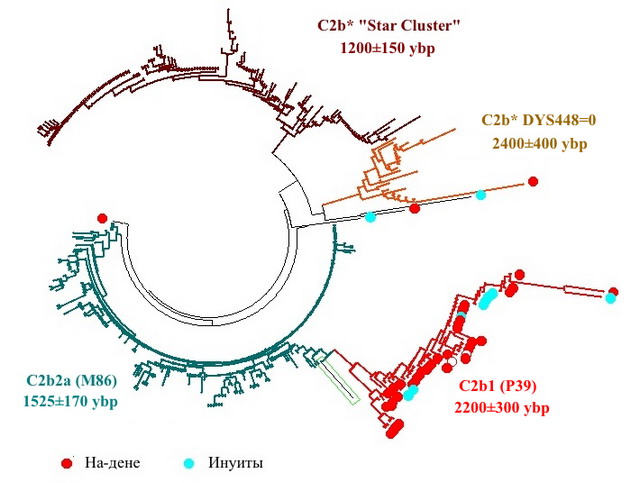
Все 4 ветви субклада C2b, что представлены на диаграмме, относительно молоды. Их базовые гаплотипы в 19-маркерном формате выглядят так:
13 25 16 10 12 13 11 14 10 13 11 29 - 18 14 22 11 15 10 21 [C2b* Star cluster]
13 24 15 10 12 15 11 13 11 13 11 29 - 17 14 0 11 17 11 21 [C2b* DYS448=0]
12 23 15 9 12 15 11 13 11 13 11 30 - 17 14 21 11 15 10 24 [C2b1]
13 24 16 9 12 12 11 13 11 14 11 31 - 18 14 20 10 15 10 23 [C2b2a]
Они практически равноудалены друг от друга, расходясь на 9-14 мутаций между собой. Это соответствует общему предку всех ветвей, жившему 8200±3000 лет назад. Даже с учетом довольно большого разброса в датировках, этот предок приходится на послеледниковую эпоху, как и предок дальневосточной и эскимосской ветвей гаплогруппы Q. Для всех этих ветвей известны также протяженные 67-маркерные базовые гаплотипы, хотя и на более ограниченных выборках. Чтобы не перегружать читателей цифрами, не буду их выписывать, а сразу перейду к датировке общего предка. Она практически совпадает с той, что была получена в коротких форматах - 7700±2000 лет назад. Наконец, счет по медленной 22-маркерной панели дает чуть более древнее время, но с очень большим разбросом - 13000±7500 лет назад.
Из всех этих расчетов можно сделать вывод, что, как и у эскимосов, у народов на-дене ведущая генеалогическая линия восходит к потомкам мигрантов из северо-восточной Азии, пересекшим Берингов пролив в послеледниковую эпоху. Маршруты и датировки этих переходов еще предстоит выяснить. Если судить по умению как эскимосов, так и индейцев Аляски и северо-запада Канады в строительстве судов для дальних морских переходов (см. в начале статьи репродукцию с гравюры 18-го века, изображающую большое каноэ тлингитов), то напрашивается водный путь.
К принципиальным результатам исследования можно отнести тот факт, что абстрактная лингвистическая конструкция о родстве языков на-дене с большой группой языков Евразии обретает вполне осязаемые корни в лице живых людей из конкретных ДНК-родов. Не должно смущать то, что специфической для на-дене оказалась гаплогруппа С, а не Q или R, что ассоциируются с дене-кавказской макросемьей. Относительно недавняя датировка общего предка ветви C2b1 (P39) предполагает, что исходная популяция, пришедшая из Азии, уже в Америке прошла через бутылочное горлышко, и выжила только одна ветвь, не обязательно основная в исходном наборе. Строго говоря, среди носителей гаплогруппы Q у на-дене имеются две небольшие по размеру ветви, что не входят в «индейский» субклад М3. Они были тестированы как Q1a* (MEH2) и Q1a2a1* (L54). Их можно было бы отнести к реликтам, оставшимся от тех пресекшихся линий, но пока недостаточно данных, чтобы это подтвердить или опровергнуть.
Игорь Рожанский, кандидат химических наук
«Переформат», 15 июня 2014Из комментариев:
Прыг-скок говорит - >> С тех пор, как дене-кавказская гипотеза была впервые сформулирована С. Старостиным в 1984 году, было собрано довольно много языкового материала, подтверждающего, по мнению ее сторонников, родство языков на-дене, по меньшей мере, с енисейскими.
Если правильно помню, то в 80ые у Старостина речь шла о родстве северокавказских, енисейских и сино-тибетских. Потом уже стали добавлять другие семьи и изоляты. Кстати, дене-енисейская гипотеза Вайды вроде бы никакого отношения к этому не имеет - самостоятельная гипотеза. По Старостину енисейские сближали с сино-тибетскими в рамках дене-кавказских - хотя там разные версии ходят.
И. Рожанский отвечает: - Для той постановки задачи, что сформулирована в заметке, гипотезы Старостина и Вайды равнозначны. Обе в неявной форме постулируют языковый контакт между жителями Сибири и Аляски в послеледниковую эпоху. Причем в концепции Э. Вайды, сторонника традиционных методов компаративистики, этот контакт должен был иметь место позднее, чем в модели убежденного ностратиста С. Старостина. Однако авторы обеих лингвистических гипотез, похоже, не особенно акцентировали внимание на механизме того, когда и каким образом языки перешли из Сибири в Америку.
Данные ДНК на-дене и эскимосов, во-первых, подтверждают факт послеледниковой миграционной волны, и, во-вторых, дают довольно широкий временной промежуток, когда она могла осуществиться. Ориентировочно, это между 10000 и 4000 лет назад для эскимосов и между 10000 и 2500 лет назад для на-дене. Остальные детали пазла под названием «дене-кавказская» или «дене-енисейская семья» в руках у археологов, антропологов, палеоклиматологов и палеоботаников.
Прыг-скок говорит: - A. Kassian. Hattic as a Sino-Caucasian language // Ugarit-Forschungen. Bd 41, 2009-2010. P. 309-447.

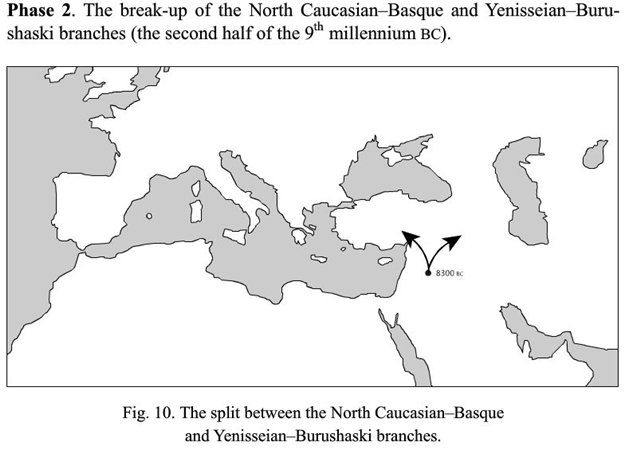
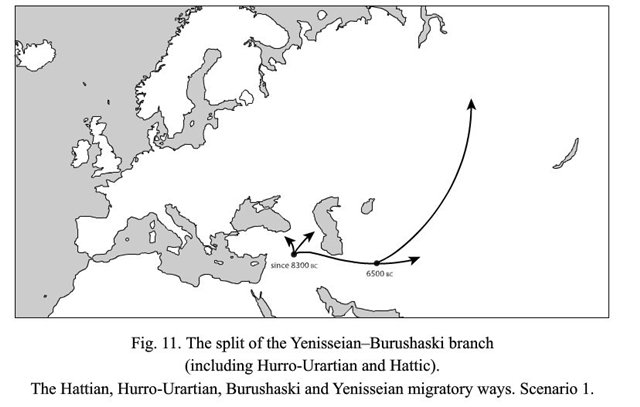

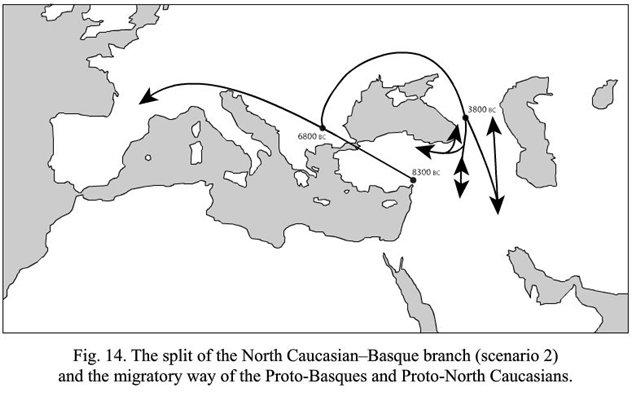
Очень интересный вариант со сближением енисейских и бурушаски с древневосточными хаттским и хурро-урартскими (последние два оказываются не близки северокавказским, сближаемым с баскским). На-дене тут с енисейскими опять-таки не сближены.
И. Рожанский отвечает: - Спасибо за ссылку! Материал интересный, но непонятно, почему все сторонники теорий дальнего родства как зачарованные помещают прародину всех макросемей в Плодородный Полумесяц. И это при том, что из всей территориально разбросанной дене-кавказской макросемьи ближневосточные корни имеют только северокавказские, васконские (баскский + вымершие языки Западной Европы) и, по предположению автора, хаттский и хуррито-урартские языки. Все остальные располагаются намного восточнее.
Вместо сложной схемы с двумя-тремя независимыми путями распространения языков в восточном направлении (совсем не стыкующуюся с реконструкциями ДНК-генеалогии, кстати), намного проще предложить одну миграционную волну из Центральной Азии на Ближний Восток, что положила начало группе языков, что объединяются под термином «эрбин». На схеме внизу они обведены синим прямоугольником.
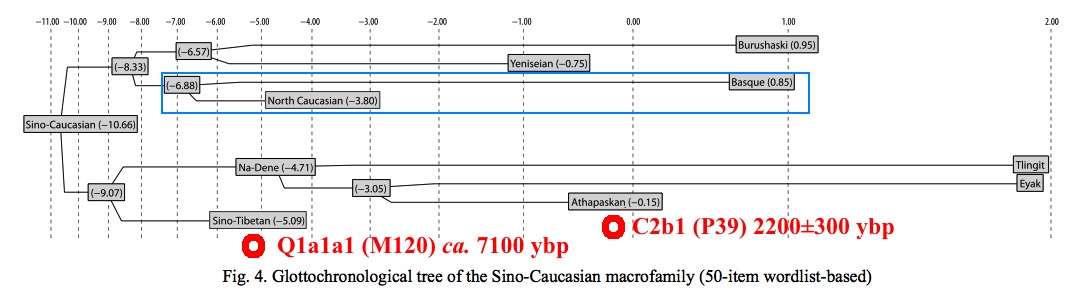
По глоттохронологическим расчетам автора, появление носителей дене-кавказских языков на Ближнем Востоке приходится на промежуток времени с 10500 до 9000 лет назад. У археологов есть разные трактовки событий, что происходили на Ближнем Востоке в те времена, но о том, что кто-то пришел с Востока, можно судить по появлению в конце этого периода технологии, незнакомой до того первым неолитийцам Леванта - керамики. То, что она была изобретена в Восточной Азии, сейчас мало у кого вызывает сомнения. Есть надежные датировки по археологическим находкам в Китае, Японии и российском Приморье, что уводят первые керамические изделия на 17-19 тысяч лет назад, когда ни о каком неолите еще и речи не было.
Вопрос заключается в том, изобрели ли керамику на Ближнем Востоке независимо или эту технологию переняли у тех, кто владел ею уже почти 10000 лет. Миграционный путь, что реконструируется по данным ДНК-генеалогии, говорит в пользу второго варианта. В пользу первого - почти религиозная вера в то, что все достижения цивилизации, в том числе и современные языки, пошли из одного центра.
Наконец, есть два примечательных совпадения в датировках глоттохронологии и ДНК-генеалогии, что отметил на схеме. Сразу же возникает вопрос - насколько корректно отнесение эскимосско-алеутстких языков к ностратической макросемье? В схеме С. Старостина они, вместе с рядом палеосибирских языков, находятся в родстве с алтайскими, но по данным ДНК-генеалогии и антропологии для их носителей реконструируется путь от Приморья вдоль тихоокеанского побережья, а это скорее говорит в пользу какой-то другой языковой семьи.
Еще раз о дене-кавказских языках: Северо-восточный след
Переселения народов / Статья 2014 года
Обсуждение гипотезы о языке «эрбин», относившимся к гипотетической же дене-кавказской макросемье, вызвало такую бурную дискуссию, что в ней стало уже сложно ориентироваться. Некоторые тезисы участников обсуждения повторяются раз за разом, и уже трудно уследить за основной мыслью. ©Ещё в цикле Народы, где ваши гены?
Большое каноэ тлингитов, репродукция с гравюры 18-го века
Постараюсь разорвать порочный круг и дополнить материал по дене-кавказской гипотезе анализом самой дальней ветви этих языков - на-дене. Точнее, их носителей, живущих на Аляске и северо-западе Канады.
С тех пор, как дене-кавказская гипотеза была впервые сформулирована С. Старостиным в 1984 году, было собрано довольно много языкового материала, подтверждающего, по мнению ее сторонников, родство языков на-дене, по меньшей мере, с енисейскими. Однако все эти аргументы повисают в воздухе, если нет информации, когда и каким образом эти языки попали в Америку. Датировки выделения ветви на-дене приходятся на времена, когда уже образовался Берингов пролив, а Америка была заселена носителями других америндских языков. Отсюда закономерно следует предположение, что предки нынешних атапасков, навахо и воспетых в вестернах апачей переселились из Азии в Америку по воде, несколько ранее или примерно в то же время, что и предки эскимосов.
Насколько мне известно, археологи не находят однозначных свидетельств такого перехода, и лингвистам приходится опираться только на данные этнографии по сходству материальной культуры народов Аляски и Сибири. Однако датировок из такого материала не извлечь, а потому независимых подтверждений о родстве языков на-дене с другими языками гипотетической макросемьи практически нет.
Популяционные генетики в своих исследования коренных народов Америки также не находят доказательств нескольких волн миграции, но их выводы были сделаны в отсутствие данных по народам на-дене, не говоря уж о, мягко говоря, спорных методах их расчетов. Недавно появилась возможность рассчитать датировки, используя данные по ДНК коренных народов Аляски и северо-западной Канады, опубликованные в статьях C. Davis с соавторами 2011 года и M. Dulik с соавторами 2012 года. Всего было опубликовано 233 Y-хромосомных гаплотипа различных племен на-дене в 17- и 19-маркерном форматах, а также 363 гаплотипа из трех этнических групп эскимосов: юпик, инувиалук и инупиат. Как это уже неоднократно наблюдалось для других групп аборигенов, многие из их представителей оказались потомками европейских поселенцев по своей прямой мужской линии. Это около половины индейцев на-дене и чуть меньше четверти эскимосов. Оставшиеся 120 «чистокровных» на-дене и 264 эскимоса распределились по гаплогруппам С и Q.
В статье Dulik с соавторами в популяции эскимосов был обнаружен новый снип гаплогруппы Q (NWT01), на основании которого авторы выделили новый субклад Q1a6 и рассчитали время жизни общего предка его носителей - около 7500 лет назад. Казалось бы, чем не датировка переселения в Америку в послеледниковую эпоху? Увы, считали с помощью «эволюционных скоростей», а потому датировка не имеет ничего общего с реальностью. Еще спустя какое-то время нашли, что новый субклад Q1a6 оказался в не слишком дальнем родстве с ветвью, имевшей тогда обозначение Q1a1 (М120). Вот как выглядит соответствующий фрагмент древа гаплогруппы Q после уточнения:
Напомню, что американские индейцы относятся к другой ветви этой гаплогруппы, а именно Q1a2a1 (L54), и их общий предок с найденной у эскимосов генеалогической линией уходит не менее чем на 20000 лет назад. Родство последней с дальневосточной ветвью М120 дает возможность предположить, что мы видим след миграции в Америку, прошедшей после того, как растаял ледник, и перешеек между Чукоткой и Аляской скрылся под водой. Чтобы проверить это предположение, надо рассчитать, как давно жил общий предок двух ветвей, во время существования Берингии или после.
Для начала рассмотрим древо 17-маркерных гаплотипов гаплогруппы Q из выборок на-дене, эскимосов и народов Дальнего Востока, представителей ветви М120.
Аляскинская NWT01 и дальневосточная М120 располагаются на диаграмме ближе друг к другу, как и следовало бы ожидать. Первая из них сходится к предку, жившему 2500±300 лет назад, и имеет такой базовый 19-маркерный гаплотип в формате YFiler, с добавленными DYS426 и DYS388:
14 24 13 10 13 19 12 12 11 14 15 30 - 14 15 18 10 15 11 22
Вторая, М120, состоит из двух подветвей. Та, что выделена на диаграмме оранжевым цветом, имеет «возраст» 4000±450 лет и базовый гаплотип:
14 24 13 9 15 22 12 12 11 14 14 30 - 16 14 19 10 16 12 22
Ветвь, выделенная коричневым, соответственно, 3700±560 лет и
14 24 13 6 16 20 12 12 11 13 14 30 - 16 14 19 10 17 12 22
С учетом того, что очень редкое значение DYS391=6 (четвертый слева маркер) во второй ветви появилось, очевидно, в результате одной многошаговой мутации, эти две ветви расходятся на 7 мутаций, что соответствует дистанции в 6450 лет. С учетом «возраста» базовых гаплотипов это дает (4000 + 3700 + 6450)/2 ≈ 7100 лет до их общего предка. Сравним теперь эти базовые гаплотипы с ветвью, найденной у эскимосов Аляски. Получаем дистанции в 13 и 16 мутаций соответственно, что при пересчете дает общего предка, жившего около 11000 или 13500 лет назад, при оценке по первой и второй ветви. В пределах погрешности для столь давних времен это одно и то же. Для сравнения, европейские и американские линии субклада Q1a2a1 (L54) разошлись около 17000 лет назад, открытие прохода в ледовом щите между Аляской и территорией современного штата Монтана датируется временем около 13000 лет назад, а возникновение Берингова пролива - 11000 лет назад.
Было бы желательно уточнить датировку общего предка, используя протяженные гаплотипы, но пока таких данных нет. На сегодняшний момент можно сделать вывод, что предок генеалогических линий субклада Q1a1a из Китая и с Аляски жил, очевидно, в Азии тогда, когда Северную и (возможно) Южную Америку уже населяли предки современных индейцев, а сухопутный проход или ушел под воду, или значительно сократился по своей площади. Следовательно, ДНК-генеалогия дает весомые аргументы в пользу того, что предки современных эскимосов мигрировали из Азии в Америку позже, чем предки американских индейцев, скорее всего, водным путем. Когда именно это произошло, ни археология, ни ДНК-генеалогия, ни, тем более, лингвистика не дают однозначного ответа.
Верхний предел можно оценить, исходя из анализа других генеалогических линий эскимосов. В исследуемой выборке к ветви Q1a1a* (NWT01) относятся 115 из 264 эскимосских гаплотипов из аборигенных ветвей, то есть 44%. Еще 139 принадлежат к субкладу Q1a2a1a1 (M3), тому, что доминирует у индейцев как Северной, так и Южной Америки, но практически не встречается в Старом Свете. Очевидно, в этногенезе эскимосов приняли участие не только люди, пересекшие Берингов пролив в пост-ледниковую эпоху, но и потомки тех, кто осваивал север Америки несколькими тысячелетиями ранее. Из нескольких ветвей субклада М3, найденных в выборке, самая большая по численности (86 гаплотипов) состоит исключительно из эскимосов. На диаграмме она занимает ее нижнюю часть. Предок этой моноэтнической ветви жил 4000±460 лет назад, а ее базовый гаплотип выглядит следующим образом:
14 24 13 10 16 18 12 12 13 14 14 30 - 17 15 21 11 16 10 22
Поиск в доступных базах данных не выявил каких-либо близких к ней гаплотипов американских индейцев. Это, скорее всего, означает, что носители этой ветви влились в состав прото-эскимосов на самом раннем этапе этногенеза. Поскольку субклад М3 по всем признакам зародился в Америке, то это означает, что самое позднее 4000 лет назад предки современных эскимосов уже жили на севере Америки. Эту датировку можно рассматривать как подсказку для археологов в их интерпретациях древних культур Арктики.
Однако пока что нет ответа на поставленную задачу - доказать или опровергнуть факт миграции предков на-дене из Азии в послеледниковую эпоху. В ветви NWT01 народы на-дене представлены единичными гаплотипами, в основной же своей массе они принадлежат к различным ветвям субклада М3, что на диаграмме располагаются в левом нижнем секторе. Как уже говорилось выше, субклад М3 характеризует практически все группы американских индейцев, и в этой части своих генеалогических линий на-дене ничем не выделяются на их фоне. Если ограничиваться анализом гаплогруппы Q, то вопрос о второй волне миграции из Азии для предков на-дене можно считать закрытым - в их Y-ДНК ее следы отсутствуют. В свою очередь, этот факт ставит под серьезное сомнение их языковое родство с енисейскими, сино-тибетскими и т.д. Даже самые изощренные методы компаративистики не в силах распознать родство языков, разошедшихся 17000 лет назад или ранее.
А если искать следы общего происхождения за пределами гаплогруппы Q? На-дене выделяются на фоне других коренных народов Америки тем, что помимо гаплогруппы Q у них в заметной пропорции представлена гаплогруппа С. В данной выборке это ровно ⅓ (40 из 120-ти) индейцев из этнических групп атабасков, в числе которых племена тличо, кучин, тлингитов и т.д. Из этих 40 человек 37 относятся к ветви, что характеризуется снипом Р39. В текущей нотации она обозначается как C2b1, и на дереве гаплогруппы С располагается в том же субкладе C2b (L1373), что и ветви, широко представляющие у монголов, казахов, киргизов и других народов Центральной Азии. Вот как выглядит их общее дерево.
Все 4 ветви субклада C2b, что представлены на диаграмме, относительно молоды. Их базовые гаплотипы в 19-маркерном формате выглядят так:
13 25 16 10 12 13 11 14 10 13 11 29 - 18 14 22 11 15 10 21 [C2b* Star cluster]
13 24 15 10 12 15 11 13 11 13 11 29 - 17 14 0 11 17 11 21 [C2b* DYS448=0]
12 23 15 9 12 15 11 13 11 13 11 30 - 17 14 21 11 15 10 24 [C2b1]
13 24 16 9 12 12 11 13 11 14 11 31 - 18 14 20 10 15 10 23 [C2b2a]
Они практически равноудалены друг от друга, расходясь на 9-14 мутаций между собой. Это соответствует общему предку всех ветвей, жившему 8200±3000 лет назад. Даже с учетом довольно большого разброса в датировках, этот предок приходится на послеледниковую эпоху, как и предок дальневосточной и эскимосской ветвей гаплогруппы Q. Для всех этих ветвей известны также протяженные 67-маркерные базовые гаплотипы, хотя и на более ограниченных выборках. Чтобы не перегружать читателей цифрами, не буду их выписывать, а сразу перейду к датировке общего предка. Она практически совпадает с той, что была получена в коротких форматах - 7700±2000 лет назад. Наконец, счет по медленной 22-маркерной панели дает чуть более древнее время, но с очень большим разбросом - 13000±7500 лет назад.
Из всех этих расчетов можно сделать вывод, что, как и у эскимосов, у народов на-дене ведущая генеалогическая линия восходит к потомкам мигрантов из северо-восточной Азии, пересекшим Берингов пролив в послеледниковую эпоху. Маршруты и датировки этих переходов еще предстоит выяснить. Если судить по умению как эскимосов, так и индейцев Аляски и северо-запада Канады в строительстве судов для дальних морских переходов (см. в начале статьи репродукцию с гравюры 18-го века, изображающую большое каноэ тлингитов), то напрашивается водный путь.
К принципиальным результатам исследования можно отнести тот факт, что абстрактная лингвистическая конструкция о родстве языков на-дене с большой группой языков Евразии обретает вполне осязаемые корни в лице живых людей из конкретных ДНК-родов. Не должно смущать то, что специфической для на-дене оказалась гаплогруппа С, а не Q или R, что ассоциируются с дене-кавказской макросемьей. Относительно недавняя датировка общего предка ветви C2b1 (P39) предполагает, что исходная популяция, пришедшая из Азии, уже в Америке прошла через бутылочное горлышко, и выжила только одна ветвь, не обязательно основная в исходном наборе. Строго говоря, среди носителей гаплогруппы Q у на-дене имеются две небольшие по размеру ветви, что не входят в «индейский» субклад М3. Они были тестированы как Q1a* (MEH2) и Q1a2a1* (L54). Их можно было бы отнести к реликтам, оставшимся от тех пресекшихся линий, но пока недостаточно данных, чтобы это подтвердить или опровергнуть.
Игорь Рожанский, кандидат химических наук
«Переформат», 15 июня 2014Из комментариев:
Прыг-скок говорит - >> С тех пор, как дене-кавказская гипотеза была впервые сформулирована С. Старостиным в 1984 году, было собрано довольно много языкового материала, подтверждающего, по мнению ее сторонников, родство языков на-дене, по меньшей мере, с енисейскими.
Если правильно помню, то в 80ые у Старостина речь шла о родстве северокавказских, енисейских и сино-тибетских. Потом уже стали добавлять другие семьи и изоляты. Кстати, дене-енисейская гипотеза Вайды вроде бы никакого отношения к этому не имеет - самостоятельная гипотеза. По Старостину енисейские сближали с сино-тибетскими в рамках дене-кавказских - хотя там разные версии ходят.
И. Рожанский отвечает: - Для той постановки задачи, что сформулирована в заметке, гипотезы Старостина и Вайды равнозначны. Обе в неявной форме постулируют языковый контакт между жителями Сибири и Аляски в послеледниковую эпоху. Причем в концепции Э. Вайды, сторонника традиционных методов компаративистики, этот контакт должен был иметь место позднее, чем в модели убежденного ностратиста С. Старостина. Однако авторы обеих лингвистических гипотез, похоже, не особенно акцентировали внимание на механизме того, когда и каким образом языки перешли из Сибири в Америку.
Данные ДНК на-дене и эскимосов, во-первых, подтверждают факт послеледниковой миграционной волны, и, во-вторых, дают довольно широкий временной промежуток, когда она могла осуществиться. Ориентировочно, это между 10000 и 4000 лет назад для эскимосов и между 10000 и 2500 лет назад для на-дене. Остальные детали пазла под названием «дене-кавказская» или «дене-енисейская семья» в руках у археологов, антропологов, палеоклиматологов и палеоботаников.
Прыг-скок говорит: - A. Kassian. Hattic as a Sino-Caucasian language // Ugarit-Forschungen. Bd 41, 2009-2010. P. 309-447.
Очень интересный вариант со сближением енисейских и бурушаски с древневосточными хаттским и хурро-урартскими (последние два оказываются не близки северокавказским, сближаемым с баскским). На-дене тут с енисейскими опять-таки не сближены.
И. Рожанский отвечает: - Спасибо за ссылку! Материал интересный, но непонятно, почему все сторонники теорий дальнего родства как зачарованные помещают прародину всех макросемей в Плодородный Полумесяц. И это при том, что из всей территориально разбросанной дене-кавказской макросемьи ближневосточные корни имеют только северокавказские, васконские (баскский + вымершие языки Западной Европы) и, по предположению автора, хаттский и хуррито-урартские языки. Все остальные располагаются намного восточнее.
Вместо сложной схемы с двумя-тремя независимыми путями распространения языков в восточном направлении (совсем не стыкующуюся с реконструкциями ДНК-генеалогии, кстати), намного проще предложить одну миграционную волну из Центральной Азии на Ближний Восток, что положила начало группе языков, что объединяются под термином «эрбин». На схеме внизу они обведены синим прямоугольником.
По глоттохронологическим расчетам автора, появление носителей дене-кавказских языков на Ближнем Востоке приходится на промежуток времени с 10500 до 9000 лет назад. У археологов есть разные трактовки событий, что происходили на Ближнем Востоке в те времена, но о том, что кто-то пришел с Востока, можно судить по появлению в конце этого периода технологии, незнакомой до того первым неолитийцам Леванта - керамики. То, что она была изобретена в Восточной Азии, сейчас мало у кого вызывает сомнения. Есть надежные датировки по археологическим находкам в Китае, Японии и российском Приморье, что уводят первые керамические изделия на 17-19 тысяч лет назад, когда ни о каком неолите еще и речи не было.
Вопрос заключается в том, изобрели ли керамику на Ближнем Востоке независимо или эту технологию переняли у тех, кто владел ею уже почти 10000 лет. Миграционный путь, что реконструируется по данным ДНК-генеалогии, говорит в пользу второго варианта. В пользу первого - почти религиозная вера в то, что все достижения цивилизации, в том числе и современные языки, пошли из одного центра.
Наконец, есть два примечательных совпадения в датировках глоттохронологии и ДНК-генеалогии, что отметил на схеме. Сразу же возникает вопрос - насколько корректно отнесение эскимосско-алеутстких языков к ностратической макросемье? В схеме С. Старостина они, вместе с рядом палеосибирских языков, находятся в родстве с алтайскими, но по данным ДНК-генеалогии и антропологии для их носителей реконструируется путь от Приморья вдоль тихоокеанского побережья, а это скорее говорит в пользу какой-то другой языковой семьи.