Про брачное доминирование у дятлов
У уток есть такой ритуал натравливания (самца самкой на «чужаков»),который у анцестральных групп не ритуализован вовсе, а затем интересным образом ритуализуется по-разному в разных ветвях семейства утиных. Классическое описание натравливания дал Лоренц («Агрессия. Так называемое «зло»):
«Прекрасный пример того, как ритуал возникает филогенетически, как он приобретает свой смысл и как изменяется в ходе дальнейшего развития, -- предоставляет нам изучение одной церемонии у самок утиных птиц, так называемого натравливания. Как и у многих других птиц с такой же семейной организацией, у уток самки хотя и меньше размером, но не менее агрессивны, чем самцы.
Поэтому при столкновении двух пар часто случается, что распаленная яростью утка продвигается к враждебной паре слишком далеко, затем пугается собственной храбрости и торопится назад, под защиту более сильного супруга. Возле него она испытывает новый прилив храбрости и снова начинает угрожать враждебной паре, но на этот раз уже не расстается с безопасной близостью своего селезня.
В своем первоначальном виде эта последовательность действий совершенно произвольна по форме, в зависимости от игры противоположных побуждений, стимулирующих утку. Временная последовательность, в которой преобладают боевой задор, страх, поиск защиты и новое стремление к нападению, легко и ясно читается по выразительным движениям утки, и прежде всего по ее положению в пространстве.
Например, у нашей европейской пеганки весь этот процесс не содержит никаких закрепленных ритуалом элементов, кроме определенного движения головы, связанного с особым звуком. Как всякая подобная ей птица, при атаке утка бежит в сторону врага, низко вытянув шею, а затем, тотчас же подняв голову, обратно к супругу. Очень часто утка, убегая, заходит за селезня и огибает его полукругом, так что в результате - когда она снова начинает угрожать -- оказывается в позиции сбоку от супруга, с головой, обращенной прямо в сторону вражеской пары.

Но часто, если бегство было не слишком паническим, она довольствуется тем, что только подбегает к своему селезню и останавливается перед ним, грудью к нему, так что для угрозы в сторону неприятеля ей приходится повернуть голову и вытянуть шею через плечо назад. Бывает и так, что она стоит боком, перед селезнем или позади него, и вытягивает шею под прямым углом к продольной оси тела, -- короче говоря, угол между продольной осью тела и вытянутой шеей зависит исключительно от того, где находится она сама, ее селезень и враг, которому она угрожает. Ни одно положение не является для нее предпочтительным.
У близкородственного огаря … натравливание уже несколько более ритуализовано. Хотя у этого вида самка "еще" может стоять рядом с супругом и угрожать прямо перед собой или, обегая вокруг него, направлять свою угрозу под любым углом к продольной оси собственного тела, -- однако в подавляющем большинстве случаев она стоит перед селезнем, грудью к нему, и угрожает через-плечо-назад. И когда я видел однажды, как утка изолированной пары этого вида производила движения натравливания "вхолостую" -- т.е. при отсутствии раздражающего объекта, - она тоже угрожала через-плечо-назад, как будто видела несуществующего врага именно в этом направлении.
У настоящих уток -- к которым принадлежит и наша кряква, предок домашней утки, -- натравливание черезплечо-назад превратилось в единственно возможную, обязательную форму движения, так что самка, прежде чем начать натравливание, всегда становится грудью к селезню, как можно ближе к нему; соответственно, когда он бежит или плывет -- она следует за ним вплотную. Интересно, что движение головы через-плечо-назад до сих пор включает в себя первоначальные ориентировочные реакции, которые у всех видов Таdornа породили фенотипически -- т.е. с точки зрения формы, внешнего облика -- подобную, но изменчивую форму движения. Лучше всего это заметно, когда утка начинает натравливание в состоянии очень слабого возбуждения и лишь постепенно приводит себя в ярость. При этом может случиться, что поначалу - если враг стоит прямо перед ней -- она станет угрожать прямо вперед; но по мере того как возрастает ее возбуждение, она проявляет неодолимое стремление вытянуть шею назад через плечо.
Что при этом всегда существует и другая ориентирующая реакция, которая стремится обратить угрозу в сторону врага, - это можно буквально "прочесть по глазам" утки: взгляд ее неизменно прикован к предмету ее ярости, хотя новая, твердо закрепленная координация движения тянет ее голову в другую сторону. Если бы утка говорила, она наверняка сказала бы: "Я хочу пригрозить вон тому ненавистному чужому селезню, но что-то оттягивает мне голову!" Наличие двух соперничающих друг с другом тенденций движения можно доказать объективно и количественно, а именно: если чужая птица, к которой обращена угроза, стоит перед уткой, то отклонение головы в сторону поворота назад является наименьшим. Оно увеличивается в точности настолько, насколько увеличивается угол между продольной осью тела утки и направлением на врага. Если он стоит прямо за нею, т.е. угол составляет 180 градусов, то утка при натравливании почти достает клювом собственный хвост [Прим. перев. - Очевидно, автор имел в виду, что, по мере нарастания возбуждения, утка сама отворачивается от "врага" и в конце концов достает клювом собственный хвост.].

Это конфликтное поведение уток при натравливании допускает лишь одно-единственное толкование, которое должно быть верным, каким бы странным оно ни казалось на первый взгляд. К легкоразличимым факторам, из которых первоначально возникли описанные движения, в ходе эволюционного развития вида присоединился еще один, новый, Как уже сказано, у пеганки бегство к супругу и нападение на врага "еще" вполне достаточны, чтобы полностью объяснить поведение утки. Совершенно очевидно, что у кряквы действуют такие же побуждения, но на обусловленные ими движения накладывается новое, независимое от них. Сложность, чрезвычайно затрудняющая анализ общей картины, состоит в том, что вновь возникшее в результате ритуализации инстинктивное действие является наследственно закрепленной копией тех действий, которые первоначально вызывались другими стимулами.
Разумеется, это действие от случая к случаю проявляется очень различно -- при различной силе вызывающих его независимых стимулов, -- так что вновь возникающая жесткая инстинктивная координация представляет собой лишь один часто встречающийся вариант. Этот вариант затем схематизируется -- способом, весьма напоминающим возникновение символов в истории человеческой культуры. У кряквы первоначальное разнообразие направлений, в которых могли находиться супруг и противник, схематически сузилось таким образом, что первый должен стоять перед уткой, а второй за нею; из агрессивного "туда" к противнику и из мотивированного бегством "сюда" к супругу получается слитое в жесткую церемонию и весьма упорядоченное "туда-сюда", в котором эта упорядоченность, регулярность уже сама по себе усиливает выразительность движений.
Вновь возникшее инстинктивное движение становится господствующим не сразу; поначалу оно всегда существует наряду с неритуализованным образцом и в первое время лишь слегка на него накладывается. Например, у огаря зачатки координации, заставляющей голову утки двигаться при натравливании назад через плечо, можно заметить лишь в том случае, если церемония выполняется "вхолостую", т.е. при отсутствии врага. В противном случае угрожающее движение обязательно направляется на него, за счет преобладания первичных направляющих механизмов».
***
Аналогичное поведение обнаруживается в совершенно другой группе, у пёстрых дятлов, рода Dendrocopos и Picoides. Формируясь на основе того же самого мотивационного конфликта, оно играет иную функциональную роль - способствует сближению партнёров в процессе ухаживания, снижая риск его срыва агрессией и уходом второй особи (что более чем существенно у столь агрессивных птиц, как дятлы), а потом, после консолидации пары, определяет характер «разделения труда» между самцом и самкой.
В отличие от уток дятлы строго моногамны, пары постоянны как минимум в репродуктивный сезон. «Натравливание» здесь включено в более сложный комплекс поведения, который может быть назван брачным доминированием (самца над самкой в паре или наоборот), а «натравливание» особей подчинённого пола на аналогичных особей из соседних пар их партнёрами или партнёршами является важной частью ухаживания.
Пёстрые дятлы не только высокоагрессивны, но и одиночно-территориальны зимой, поэтому главной проблемой сближения потенциальных партнёров, а потом и «законных супругов» в образовавшейся паре являются периодические всплески агрессии и попытки ухода, «прорывающиеся» из-за того, что партнёр воспринимается как потенциальный захватчик. Успешная агрессия «натравленных» птиц подчинённого пола против таких же соседей вносит в уменьшение вероятности «срыва» ухаживательного сближения самца и самки не меньший вклад, чем успех предыдущего токования сам по себе, а в период консолидации пары, выбора места для гнезда и строительства оного в апреле-начале мая - и больший.
Брачное доминирование непосредственно проявляется в способности птиц доминирующего пола (в условиях, когда самец и самка в образовавшейся паре находятся рядом и кормятся в паузе между токованиями или позже, когда они уже синхронизировали активность и перемещаются / кормятся вместе) беспрепятственно сгонять птиц подчинённого пола с мест, где они отдыхают/кормятся и начать кормиться там самому, при частом копировании кормового метода согнанного партнёра. Непосредственно после «сгона» у подчинённой особи повышается агрессивность, находящая выход через интенсификацию видоспецифичной территориальной активности-маркировок, патрулирований и конфликтов, причём «натравленные» особи активно ищут соседей, «чтобы» атаковать их при первом подозрении на нарушение границ или демаркировать последние демонстративным полётом, если сосед благоразумно держится на своей территории.
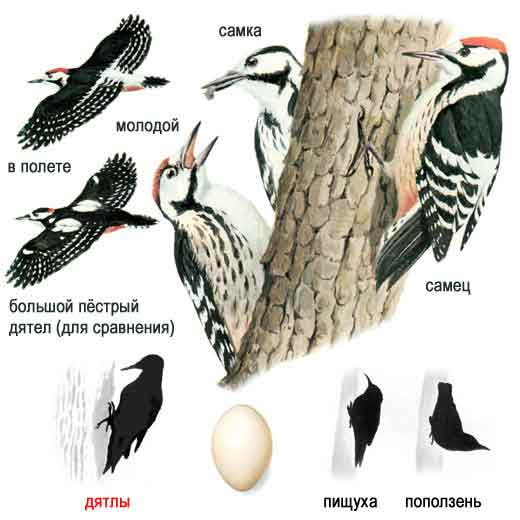
У части видов при таком доминировании на местах кормления и/или отдыха самка постоянно сгоняет самца, но не наоборот и, соответственно, доминирует над ним. Это большой пёстрый и сирийский дятлы: самки здесь немного крупнее и много агрессивнее самцов. У другой группы видов - белоспинный, трёхпалый и малый пёстрый дятлы - напротив, самцы сгоняют самок (в парах малого дятла доминирование самца частичное, он сгоняет самку примерно в 70% случаев, в остальных - она его).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Брачные пары у пёстрых дятлов могут образовываться осенью или весной. Первое преимущественно у видов с многолетне-постоянными парами (белоспинный и трёхпалый дятлы, большая часть популяции малых пёстрых дятлов, и 10-15% популяции у прочих видов. Второе типично для большей части популяции тех видов, у которых в конце сезона размножения пары вновь распадаются и живут на своих одиночных территориях осень и зиму (молодые, понятное дело, диспергируют, также как часть резидентов), а весной воссоединяются вновь, часто в прежнем составе. Это большой пёстрый, сирийский, малый и большой острокрылый дятлы.
{kind=link}
{kind=link}
[средний дятел стоит наособицу от обоих групп: его территориальное поведение редуцировано полностью, птицы не охраняют даже микротерриторий вокруг себя, поэтому сближение потенциальных партнёров не сопровождается агрессией вообще, если только рядом нет третьей особи. Но зато и пары существенно менее устойчивы - до момента окончательного выбора самцы и самки меняют по 3-5 потенциальных партнёров, и лишь образовавшиеся пары привязываются к постоянной территории, подыскивают там место для гнезда и др.). За этим исключением, причём вторичного характера, образование пар пёстрых дятлов всегда идёт соединением одиночных территорий партнёров в общую территорию пары, которую одна из этих птиц, именно, подчинённого пола, начинает охранять от соседних владельцев, а роль доминирующего пола ограничена агрессивными актами, побуждающими интенсифицировать охрану, патрулирование и т.л. ]
{kind=link}
У острокрылых дятлов и среднего дятла подобных «сгонов» не зафиксировано. Соответственно, брачное доминирование отсутствует (первично в первом и вторично во втором случае, но это отдельная история), не найдено и то «разделение труда» между самцом и самкой в процессе гнездостроения, насиживания и выкармливания, которое .описано ниже.
Эти периодические «сгоны» особей подчинённого пола полом доминирующим оказывают на него неожиданный направляющий эффект. Индуцируемая террирориальная агрессия в отношении соседей соответствующего пола резко усиливается, если те не просто находятся рядом, а ещё и токуют, так что у наиболее успешных в этом плане пар брачная активность соседей избирательно подавляется.
Если территориальная агрессия «натравленной» особи против соседних резидентов и/или «чужаков» соответствующего пола успешна, это
- сокращает срок до следующего токования с доминирующей самкой или самцом, увеличивает доступность их для ухаживания, снижает риск «выплесков» агрессии со стороны доминанта при обмене ухаживательными демонстрациями, так же как вероятность ухода и срыва взаимодействия со стороны подчинённого в ответ на агрессию;
- уменьшает страх самца и самки друг перед другом (что хорошо видно по разной частоте смещённых движений типа вышеописанных Лоренцом после успешных и неуспешных «натравливаний», в парах с высокой и низкой частотой брачного доминирования и т.д.), и
- увеличивает тяготение партнёров друг к другу в следующий раз, готовность на крик/барабанную дробь немедленно прилетать для токования, координировать перемещения и т.д.
Таким образом, успешное осуществление взаимодействий брачного доминирования переадресует взаимную агрессию партнёров на «чужаков». Это даёт возможность максимально безболезненно перейти от этапа одиночного существования, когда «следует» атаковать всех птиц, появившихся на твоём индивидуальном участке, к существованию в паре, когда для устойчивости брачных связей столь же императивно «требуется» координировать поведение, синхронизировать перемещение и т.д. с одной из таких особей.
Самое интересное, что дифференциальный успех размножения пары (предположительно) и относительная скорость прохождения парой основных стадий репродуктивного цикла (точно) от этой безболезненности зависит намного сильней, чем от качества местообитания, обилия корма и других характеристик среды - как минимум потому, что птицы не занимают территории там, где не могут обеспечить на них заведомый избыток ресурсов.
Соответственно, в период весеннего возбуждения у видов с доминированием самки у границы территорий соседние самцы конкурируют за самок и сталкиваются между собой, а она пассивно ждёт победителя, у видов с доминирование самца напротив - самки конкурируют и дерутся друг с другом за «равнодушных» самцов. На следующем этапе консолидации, после образования пары, самка (самец), конечно, не оставят партнёра (-ршу), если те проиграют стычку с соседом, но следующее сближение для ухаживания затруднится.
В результате пары, где связка «натравливание → победоносный конфликт → успешное токование» эффективно работает в каждом звене, существенно «вырываются вперёд» по сравнению со средним по поселению, раньше заканчивают строительство гнезда, интенсивнее спариваются и раньше начинают яйцекладку, а подобный сдвиг фенодат почти всегда означает больший репродуктивный успех.
Из этого распределения ролей доминирующего и подчинённого пола в процессе образования пары у всех перечисленных видов разделение функций самца и самки в последующей репродукции. У видов с доминированием самок дупло преимущественно долбят самцы, они же насиживают больше, в основном они насиживают ночью; усилия по выкармливанию обычно делятся поровну, но самка чаше «дезертирует» за несколько дней до вылета птенцов, оставляя самцу их докармливать. Слётков докармливает также преимущественно самец, что может растягиваться на одну-две недели у большого пёстрого дятла, у других видов и больше. Кроме того, самки у этих видов в начале насиживания ночуют не в гнездовом дупле, а в ночёвочном на своей бывщей зимней территории.
У видов с доминирование самцов у старых птиц (после первого размножения) всё в точности наоборот - дупло преимущественно долбит самка, она же больше насиживает, а «дезертирует» в основном самец (у малого пёстрого дятла, в известном смысле занимающего промежуточную позицию между двумя группами, такое характерно где-то для 2/3 пар). Пары из молодых птиц или пары, где один из партнёров - первогодок, показывают «разделение труда», существенно более близкое к предыдущей группе, где основные усилия по гнездостроению, насиживанию и выкармливанию возлагаются на самца (хотя система брачного доминирования в процессе ухаживания у них функционирует так же, как у взрослых). Видимо, это связано с тем, что кормодобывание молодых птиц существенно менее эффективно, чем у взрослых, и самкам нужно больше времени, чтобы набрать питательных веществ для формирования яиц.