Введение: что такое аутосомные и модальные компоненты.
Для понимая этих статей нужно понимать что такое аутосомные компоненты. Аутосомный компонент это часть статистически значимых SNP-мутаций расположенных в неполовых (то есть не в XY) хромосомах называемых аутосомами. У каждого человека есть его собственный аутосомный компонент равный его аутосомам. Любой аутосомный компонент можно разбить на несколько предковых аутосомных компонентов, которые достались ему от разных предков разной исторической глубины.
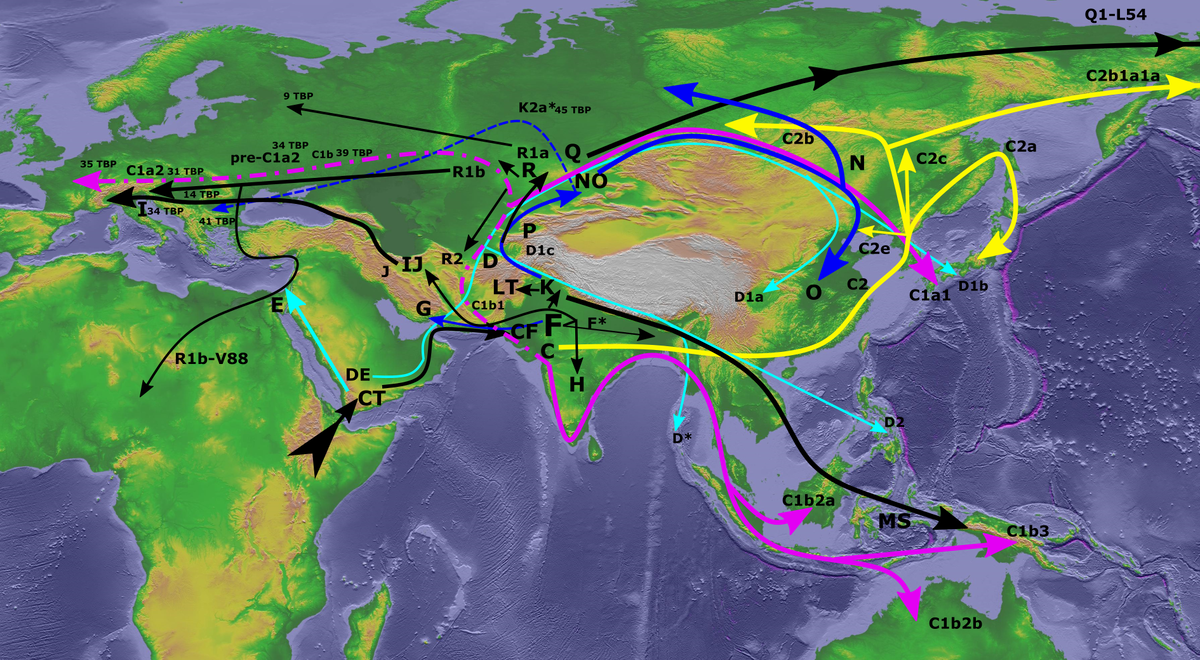
Модальный компонент это предковый аутосомный компонент связанный с его предковой Y-гаплогруппой. В истории предков данного человека он был всегда, ведь N-раз прадедушка, у которого возникла данная Y-гаплогруппа, обязательно его имел, а данный человек является его непрерывным потомком. Следовательно, его предки гарантированно его имели, что дает что дерево модальных компонентов должно совпадать с деревом Y-гаплогрупп, которое обычно формируется организацией ISOGG или компанией YFull. Приблизительное гипотетическое место возникновения Y-гаплогрупп показано на следующей карте из Википедии, в которой использована гипотеза однократного выхода кроманьонца из Африки, существует другая версия о двухкратном выходе кроманьонцев из Африки, по ней Y-гаплогруппа CТ разделилась еще в Африке, Y-гаплогруппа DE долго обитает в Африке и покидает ее через Суэц, но данная версия не учитывает обратные миграции с Ближнего Востока в Африку.
Постепенно модальный компонент меняется в результате генетического дрейфа и в результате притока других компонентов связанных с другими популяциями переданные посредством браков, поэтому у каждой гаплогруппы являющимся субкладом данной гаплогруппы он может несколько отличаться или даже полностью исчезнуть. Исчезновение модального компонента может произойти за 200 лет если человек попал в полностью другую популяцию, тогда его аутосомы полностью исчезнут за 6-7 поколений, но в глубокой древности такое событие было практически невероятным, ведь человек не мог переехать куда-то жить в одиночку, если происходили миграции, то они происходили коллективно и на относительно небольшие расстояния, где жили люди с не сильно отличающимся компонентами (конечно если это не было границей разных компонентов) и людей было мало чтобы одна популяция быстро бесследно растворилась в другой, поэтому сразу бесследно раствориться он не мог.
Модальный компонент выделяется только из палеогенетических исследований. Это компонент который найден для образцов с определенной Y-гаплогруппой. Так как мутации у Y-гаплогрупп происходят часто, раз в 100-150 лет, и в популяции в древности был небольшой набор Y-гаплогрупп, то каждый предковый аутосомный компонент можно связать с какой-то Y-гаплогруппой (возможно не одной), но нельзя связать с mt-гаплогруппой, поскольку у них мутации происходят раз 3-5 тысяч лет и они распространены на огромные растояния, к тому же женщины постоянно меняют свои рода и популяции.
Перечислим основные выделенные предковые компоненты с их описанием, кому они являются модальными и кто их образцовые носители.
Базальный компонент - виртуальный компонент, по определению возникший в Евразии до смешения кроманьонцев с неандертальцами и денисовцами. Никаких образцов его не найдено, поэтому строится только гипотетически. Очевидно был модальным компонентом для Y-гаплогруппы CТ. В моделях для него как прокси часто выступает Levant_Natufian.
ANE - древние северные евразийцы, является модальным компонентом для Y-гаплогруппы R, выделен из среднепалеолитеского мальчика R* из Мальты, Прибайкалье. Его образцовым носителем является RUS_MA1_MUP:MA1. Гораздо более молодыми представителем этого компонента являются среднепалеолитические афонтовогорцы более западные чем Мальтнец, с Енисея, его образцовым носителем является RUS_AfontovaGora3_LUP:AfontovaGora3, которые по всей видимости являются модальными для Q-F746 и R1.
ANS - древние северные сибиряки, является модальным компонентом для Y-гаплогруппы P1, выделен из среднепалеолитической стоянки в северной Сибири на реке Яна. Его образцовым носителями являются RUS_Yana_MUP. Этот потомком является ANE.
EHG - восточноевропейские охотники-собиратели, является модальным компонентом для Y-гаплогруппы R1a, выделен из мезолитических образцов Онежской культуры на Малом Оленьем Острове (Карелия, Онежское озеро), к нему относятся и первые мезолитчки Европы из культуры Веретье, самые древние образцы с этим компонентом и этой гаплогруппой. Его образцовым носителями являются RUS_Karelia_EHG. Этот компонент является потомком ANE в его позднейшей версии.
WHG - западноевропейские охотники-собиратели, вполне вероятно что является модальным компонентом для Y-гаплогруппы R1b,или тесно связан с ним, точно был модальным для R1b-V88, выделен из мезолитического образца Лошбур. Его образцовыми носителями являются из образцы Лошбура и Rochedane. Более ранняя версия этого компонента без влияние на него EHG из позднепалеолитическое эпиграветтской стоянки Виллабруна, Италия ITA_Villabruna_LUP с R1b. В предшествующий период в Европе проживали другие компоненты, которые обычно компонентами не называются, а называются кластерами по образцовым стоянкам, такое наименование вызвано тем что к современным популяциям они не попали (вымерли), и могут быть интересны только в вопросе их участия в формирования WHG.
CHG - кавказские охотники-собиратели, является модальным компонентом для субкладов Y-гаплогруппы J, выделен из образцов эпипалеолитического Сатсурблия и мезолитического Котиас, которые и являются его образцовыми носителями. Испытывал влияние EHG.
EIN или IRN - ранние иранские неолитчики, является производным от CHG в наиболее древнейшей его стадии еще без влияния EHG с большим количеством базального компонента, испытал сильно воздействие ANE. Выделен из ранних иранских неоличиков из Ганджи Дерех с R2a. Является модальным компонентом для Y-гаплогруппы R2a. Его образцовыми носителями являются из образцы IRN_Ganj_Dareh_N с R2a.
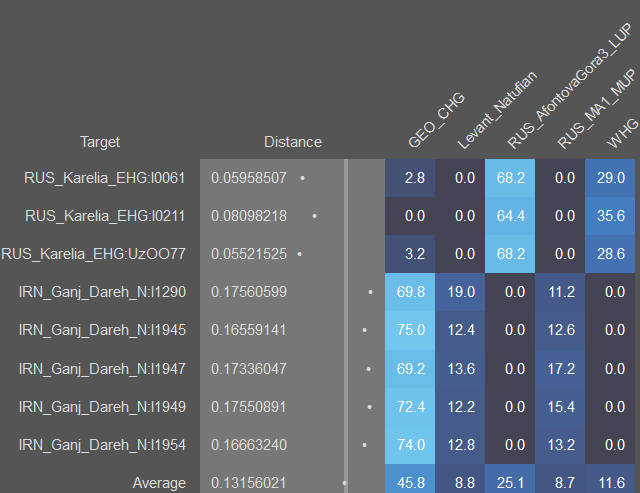
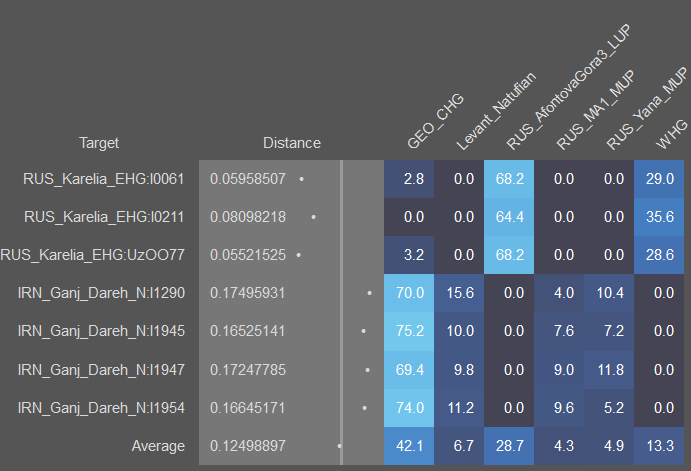
SHG - скандинавские охотники-собиратели, является суммой 40% EHG и 60% WHG. Не является модальным ни для одной гаплоруппы, поскольку чисто скандинавских мезолитических гаплогрупп не сохранилось. Его образцовыми носителями являются из образцы SWE_Motala_HG.
ANF - анатолийские неолитические фермеры, вполне вероятно что является модальным компонентом для Y-гаплогруппы G2a, но изначальных носителей которой еще не найдены. Его образцовыми носителями часто являются западноанатолийский TUR_Barcin_N, который всегда используется для рассмотрения происхождения европейских неолитических фермеров.
EEF - ранние европейские фермеры, производный от ANF. Выделяется условно по традиции, поскольку это традиционный компонент выделенный одним из первых. Вообще говоря постоянно смешивается с WHG, поэтому образцовыми для него не является никто. Поскольку у европейских неолитических фермеров был целый спектр гаплогрупп, то может быть модальным только для позднейших гаплогрупп возникших уже в неолитической Европе.
WSHG - западносибирские охотники-собиратели, позднейший охотников-собирателей из поздненеолитического времени. Сильно сдвинут к Восточной Азии по сравнению ANE афонтовогорцев. Должен быть модальным к каким-то субкладам Y-гаплогруппы Q, однако он выделен из женских образцов, поэтому его модальный компонент не определен. Его образцовым носителями являются RUS_Tyumen_WSHG и RUS_Sosnoviy_WSHG.
WSH - западные степные пастухи, также называемый степным компонентом, является суммой 60% EHG и 40% CHG. Выделен из самарских ямников. Является модальным компонентом для Y-гаплогрупп R1a-M417 и R1b-Z2103. Появился как результат смешения мужчин из Волго-Донского региона с кавказскими женщинами. Переход R1b из WHG в EHG является примером древнего эффекта основателя при попадании носителя субклада R1b в ареал EHG, в западной степи проходила граница между двумя этими компонентами. Его образцовым носителями являются Yamnaya_RUS_Samara
Существуют древние компоненты не получившие своей аббревиатуры потому что их нет в современных популяциях. Они используются по своим древним образцам. Таким является самый старый из известных человеческих образцов относящийся к инициальному верхнему палеолиту из западносибирской стоянки Усть-Ишим RUS_Ust_Ishim_IUP, он был распространен в это время в Сибири и Европе, который является модальным для Y-гаплогруппы K2a, и по всей видимости для более ранних K2, K, F и производной от нее IJ. По всей видимости был распространен в Центральной Азии и в более позднее время.
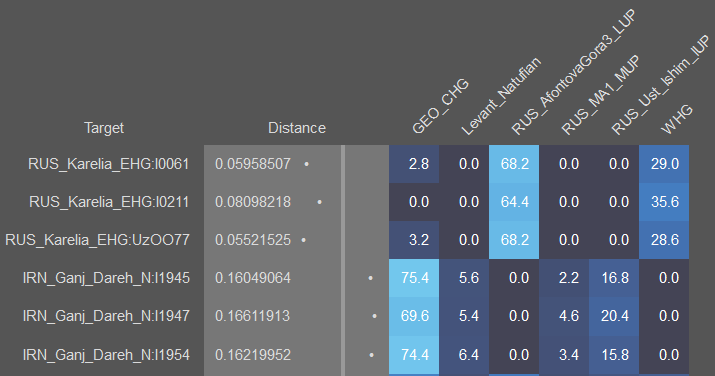
Компоненты не имеющие древних образцов тоже выделяются, они выделяются из современных представителей и называются по тем популяциям из которых они выделены. Они не являются модальными. Самыми известными такими компонентами являются нганасанский и ханьский компоненты.
ASI - древние южные индийцы, выделяется гипотетически, традиционное название для популяций современных южных индийцев без арийского влияния. Для палеообразцов придумано название AASI, но ни одного такого образца не существует, поэтому существует только в виде аббревиатуры.
Поскольку далеко не для каждого предкового аутосомного компонента существует палеообразцы, можно использовать модальные гаплогруппы в качестве нулевой гипотезы о родстве. С практической точки зрения это означает что граф адмикстур (аутосомных компонентов) должен содержать остов дерева Y-гпалогрупп к которым примешиваются в той или иной пропорции примеси из других ветвей дерева. Правильный граф адмикстур должен содержать в себе остов дерева Y-гаплогрупп, именно такие модели больше всего получаются соответствующими действительности.
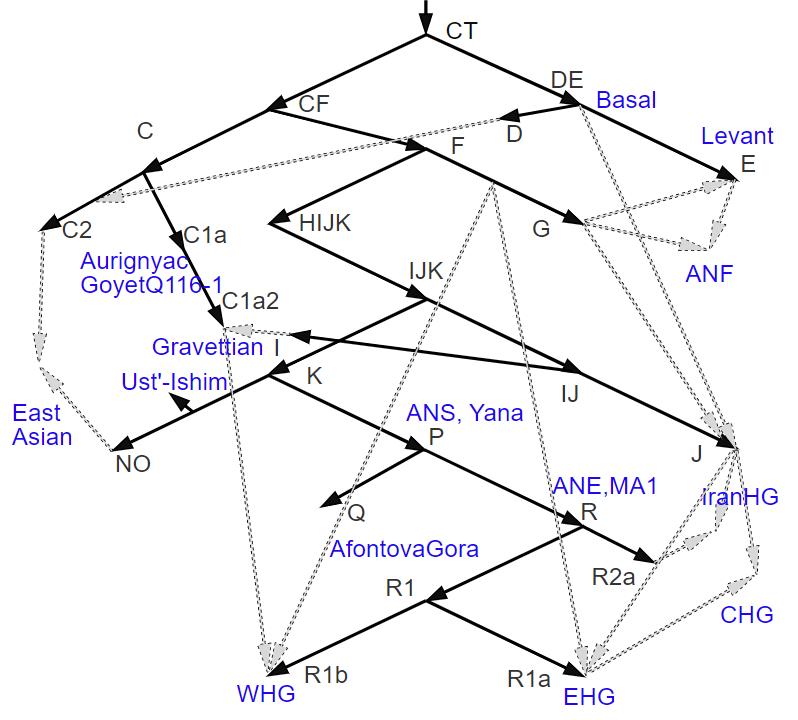
Сильно упрощенный граф на основе сильно упрощенного остовного дерева Y-гаплогрупп составленное на основе потоков из Allentoft et al. 2022..
Действительно, в программе qpGraph человек как бы сам рисует граф, а программа рассчитывает для него коэффициенты и выдает некий индекс правдоподобности. Для выбора близкого к действительности графа нужно нарисовать десятки, а то и сотни графов. Понятно, что они рисуются по представлениям человека и содержат массу ошибок, ведь вариантов множество, а человек животное ошибающееся. Вот один из таких графов из последнего препринта Allentoft et al. POPULATION GENOMICS OF STONE AGE EURASIA с добавленными мной Y-гаплогруппами.
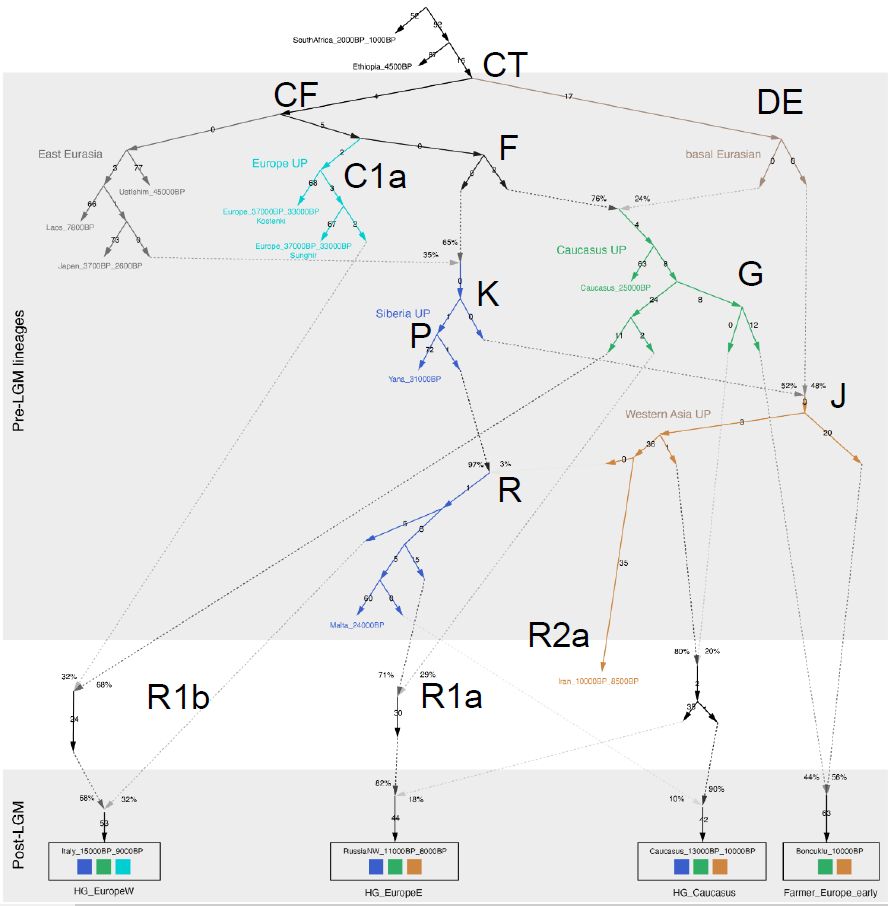
Allentoft et al.2022.
Видно что он содержит в себе некий остов дерева Y-гаплогрупп, так получилось у них в результате массового генерирования разных вариантов графа, но на нем масса ошибок связанных с неверной интерпретацией данных. Усть-Ишим у них оказался в Восточной Евразии, в то время как в нем нет ничего восточноазиатского, он существовал до геномного разделения на восточную и западную Евразию. А вот C1a у них попал в Западную Евразию, в то время как аутосомно наиболее близкий к ориньякцу ГоетуQ116-2 образец из северовосточного Китая того же времени Тяньюань, у которого есть восточноазиатский сдвиг. Тоже самое касается перевернутых стрелок, некоторые потоки шли в обратную, например в Иран был поток из ANE (R2a), но не к ANE (R).
Надо сказать что рассчитать такой граф как показано на верхнем рисунке сейчас вряд ли возможно, он слишком сложный, в нем не хватает множество еще не открытых или неопубликованных образцов. ведь на графе Алентофта есть например еще неопубликованный образец с Кавказа верхнего палеолита из Котиас, по которому есть надежда о его скорой публикации, в отличии от другого образца с Кавказа верхнего палеолита из Дзудзуаны, по которому нет надежды.
Модальный компонент это предковый аутосомный компонент связанный с его предковой Y-гаплогруппой. В истории предков данного человека он был всегда, ведь N-раз прадедушка, у которого возникла данная Y-гаплогруппа, обязательно его имел, а данный человек является его непрерывным потомком. Следовательно, его предки гарантированно его имели, что дает что дерево модальных компонентов должно совпадать с деревом Y-гаплогрупп, которое обычно формируется организацией ISOGG или компанией YFull. Приблизительное гипотетическое место возникновения Y-гаплогрупп показано на следующей карте из Википедии, в которой использована гипотеза однократного выхода кроманьонца из Африки, существует другая версия о двухкратном выходе кроманьонцев из Африки, по ней Y-гаплогруппа CТ разделилась еще в Африке, Y-гаплогруппа DE долго обитает в Африке и покидает ее через Суэц, но данная версия не учитывает обратные миграции с Ближнего Востока в Африку.
Постепенно модальный компонент меняется в результате генетического дрейфа и в результате притока других компонентов связанных с другими популяциями переданные посредством браков, поэтому у каждой гаплогруппы являющимся субкладом данной гаплогруппы он может несколько отличаться или даже полностью исчезнуть. Исчезновение модального компонента может произойти за 200 лет если человек попал в полностью другую популяцию, тогда его аутосомы полностью исчезнут за 6-7 поколений, но в глубокой древности такое событие было практически невероятным, ведь человек не мог переехать куда-то жить в одиночку, если происходили миграции, то они происходили коллективно и на относительно небольшие расстояния, где жили люди с не сильно отличающимся компонентами (конечно если это не было границей разных компонентов) и людей было мало чтобы одна популяция быстро бесследно растворилась в другой, поэтому сразу бесследно раствориться он не мог.
Модальный компонент выделяется только из палеогенетических исследований. Это компонент который найден для образцов с определенной Y-гаплогруппой. Так как мутации у Y-гаплогрупп происходят часто, раз в 100-150 лет, и в популяции в древности был небольшой набор Y-гаплогрупп, то каждый предковый аутосомный компонент можно связать с какой-то Y-гаплогруппой (возможно не одной), но нельзя связать с mt-гаплогруппой, поскольку у них мутации происходят раз 3-5 тысяч лет и они распространены на огромные растояния, к тому же женщины постоянно меняют свои рода и популяции.
Перечислим основные выделенные предковые компоненты с их описанием, кому они являются модальными и кто их образцовые носители.
Базальный компонент - виртуальный компонент, по определению возникший в Евразии до смешения кроманьонцев с неандертальцами и денисовцами. Никаких образцов его не найдено, поэтому строится только гипотетически. Очевидно был модальным компонентом для Y-гаплогруппы CТ. В моделях для него как прокси часто выступает Levant_Natufian.
ANE - древние северные евразийцы, является модальным компонентом для Y-гаплогруппы R, выделен из среднепалеолитеского мальчика R* из Мальты, Прибайкалье. Его образцовым носителем является RUS_MA1_MUP:MA1. Гораздо более молодыми представителем этого компонента являются среднепалеолитические афонтовогорцы более западные чем Мальтнец, с Енисея, его образцовым носителем является RUS_AfontovaGora3_LUP:AfontovaGora3, которые по всей видимости являются модальными для Q-F746 и R1.
ANS - древние северные сибиряки, является модальным компонентом для Y-гаплогруппы P1, выделен из среднепалеолитической стоянки в северной Сибири на реке Яна. Его образцовым носителями являются RUS_Yana_MUP. Этот потомком является ANE.
EHG - восточноевропейские охотники-собиратели, является модальным компонентом для Y-гаплогруппы R1a, выделен из мезолитических образцов Онежской культуры на Малом Оленьем Острове (Карелия, Онежское озеро), к нему относятся и первые мезолитчки Европы из культуры Веретье, самые древние образцы с этим компонентом и этой гаплогруппой. Его образцовым носителями являются RUS_Karelia_EHG. Этот компонент является потомком ANE в его позднейшей версии.
WHG - западноевропейские охотники-собиратели, вполне вероятно что является модальным компонентом для Y-гаплогруппы R1b,или тесно связан с ним, точно был модальным для R1b-V88, выделен из мезолитического образца Лошбур. Его образцовыми носителями являются из образцы Лошбура и Rochedane. Более ранняя версия этого компонента без влияние на него EHG из позднепалеолитическое эпиграветтской стоянки Виллабруна, Италия ITA_Villabruna_LUP с R1b. В предшествующий период в Европе проживали другие компоненты, которые обычно компонентами не называются, а называются кластерами по образцовым стоянкам, такое наименование вызвано тем что к современным популяциям они не попали (вымерли), и могут быть интересны только в вопросе их участия в формирования WHG.
CHG - кавказские охотники-собиратели, является модальным компонентом для субкладов Y-гаплогруппы J, выделен из образцов эпипалеолитического Сатсурблия и мезолитического Котиас, которые и являются его образцовыми носителями. Испытывал влияние EHG.
EIN или IRN - ранние иранские неолитчики, является производным от CHG в наиболее древнейшей его стадии еще без влияния EHG с большим количеством базального компонента, испытал сильно воздействие ANE. Выделен из ранних иранских неоличиков из Ганджи Дерех с R2a. Является модальным компонентом для Y-гаплогруппы R2a. Его образцовыми носителями являются из образцы IRN_Ganj_Dareh_N с R2a.
SHG - скандинавские охотники-собиратели, является суммой 40% EHG и 60% WHG. Не является модальным ни для одной гаплоруппы, поскольку чисто скандинавских мезолитических гаплогрупп не сохранилось. Его образцовыми носителями являются из образцы SWE_Motala_HG.
ANF - анатолийские неолитические фермеры, вполне вероятно что является модальным компонентом для Y-гаплогруппы G2a, но изначальных носителей которой еще не найдены. Его образцовыми носителями часто являются западноанатолийский TUR_Barcin_N, который всегда используется для рассмотрения происхождения европейских неолитических фермеров.
EEF - ранние европейские фермеры, производный от ANF. Выделяется условно по традиции, поскольку это традиционный компонент выделенный одним из первых. Вообще говоря постоянно смешивается с WHG, поэтому образцовыми для него не является никто. Поскольку у европейских неолитических фермеров был целый спектр гаплогрупп, то может быть модальным только для позднейших гаплогрупп возникших уже в неолитической Европе.
WSHG - западносибирские охотники-собиратели, позднейший охотников-собирателей из поздненеолитического времени. Сильно сдвинут к Восточной Азии по сравнению ANE афонтовогорцев. Должен быть модальным к каким-то субкладам Y-гаплогруппы Q, однако он выделен из женских образцов, поэтому его модальный компонент не определен. Его образцовым носителями являются RUS_Tyumen_WSHG и RUS_Sosnoviy_WSHG.
WSH - западные степные пастухи, также называемый степным компонентом, является суммой 60% EHG и 40% CHG. Выделен из самарских ямников. Является модальным компонентом для Y-гаплогрупп R1a-M417 и R1b-Z2103. Появился как результат смешения мужчин из Волго-Донского региона с кавказскими женщинами. Переход R1b из WHG в EHG является примером древнего эффекта основателя при попадании носителя субклада R1b в ареал EHG, в западной степи проходила граница между двумя этими компонентами. Его образцовым носителями являются Yamnaya_RUS_Samara
Существуют древние компоненты не получившие своей аббревиатуры потому что их нет в современных популяциях. Они используются по своим древним образцам. Таким является самый старый из известных человеческих образцов относящийся к инициальному верхнему палеолиту из западносибирской стоянки Усть-Ишим RUS_Ust_Ishim_IUP, он был распространен в это время в Сибири и Европе, который является модальным для Y-гаплогруппы K2a, и по всей видимости для более ранних K2, K, F и производной от нее IJ. По всей видимости был распространен в Центральной Азии и в более позднее время.
Компоненты не имеющие древних образцов тоже выделяются, они выделяются из современных представителей и называются по тем популяциям из которых они выделены. Они не являются модальными. Самыми известными такими компонентами являются нганасанский и ханьский компоненты.
ASI - древние южные индийцы, выделяется гипотетически, традиционное название для популяций современных южных индийцев без арийского влияния. Для палеообразцов придумано название AASI, но ни одного такого образца не существует, поэтому существует только в виде аббревиатуры.
Поскольку далеко не для каждого предкового аутосомного компонента существует палеообразцы, можно использовать модальные гаплогруппы в качестве нулевой гипотезы о родстве. С практической точки зрения это означает что граф адмикстур (аутосомных компонентов) должен содержать остов дерева Y-гпалогрупп к которым примешиваются в той или иной пропорции примеси из других ветвей дерева. Правильный граф адмикстур должен содержать в себе остов дерева Y-гаплогрупп, именно такие модели больше всего получаются соответствующими действительности.
Сильно упрощенный граф на основе сильно упрощенного остовного дерева Y-гаплогрупп составленное на основе потоков из Allentoft et al. 2022..
Действительно, в программе qpGraph человек как бы сам рисует граф, а программа рассчитывает для него коэффициенты и выдает некий индекс правдоподобности. Для выбора близкого к действительности графа нужно нарисовать десятки, а то и сотни графов. Понятно, что они рисуются по представлениям человека и содержат массу ошибок, ведь вариантов множество, а человек животное ошибающееся. Вот один из таких графов из последнего препринта Allentoft et al. POPULATION GENOMICS OF STONE AGE EURASIA с добавленными мной Y-гаплогруппами.
Allentoft et al.2022.
Видно что он содержит в себе некий остов дерева Y-гаплогрупп, так получилось у них в результате массового генерирования разных вариантов графа, но на нем масса ошибок связанных с неверной интерпретацией данных. Усть-Ишим у них оказался в Восточной Евразии, в то время как в нем нет ничего восточноазиатского, он существовал до геномного разделения на восточную и западную Евразию. А вот C1a у них попал в Западную Евразию, в то время как аутосомно наиболее близкий к ориньякцу ГоетуQ116-2 образец из северовосточного Китая того же времени Тяньюань, у которого есть восточноазиатский сдвиг. Тоже самое касается перевернутых стрелок, некоторые потоки шли в обратную, например в Иран был поток из ANE (R2a), но не к ANE (R).
Надо сказать что рассчитать такой граф как показано на верхнем рисунке сейчас вряд ли возможно, он слишком сложный, в нем не хватает множество еще не открытых или неопубликованных образцов. ведь на графе Алентофта есть например еще неопубликованный образец с Кавказа верхнего палеолита из Котиас, по которому есть надежда о его скорой публикации, в отличии от другого образца с Кавказа верхнего палеолита из Дзудзуаны, по которому нет надежды.