Экосистемные последствия лесоэксплуатации: Восточная Финляндия vs Карелия
Оригинал взят у wolf_kitses в Экосистемные последствия лесоэксплуатации: Восточная Финляндия vs Карелия
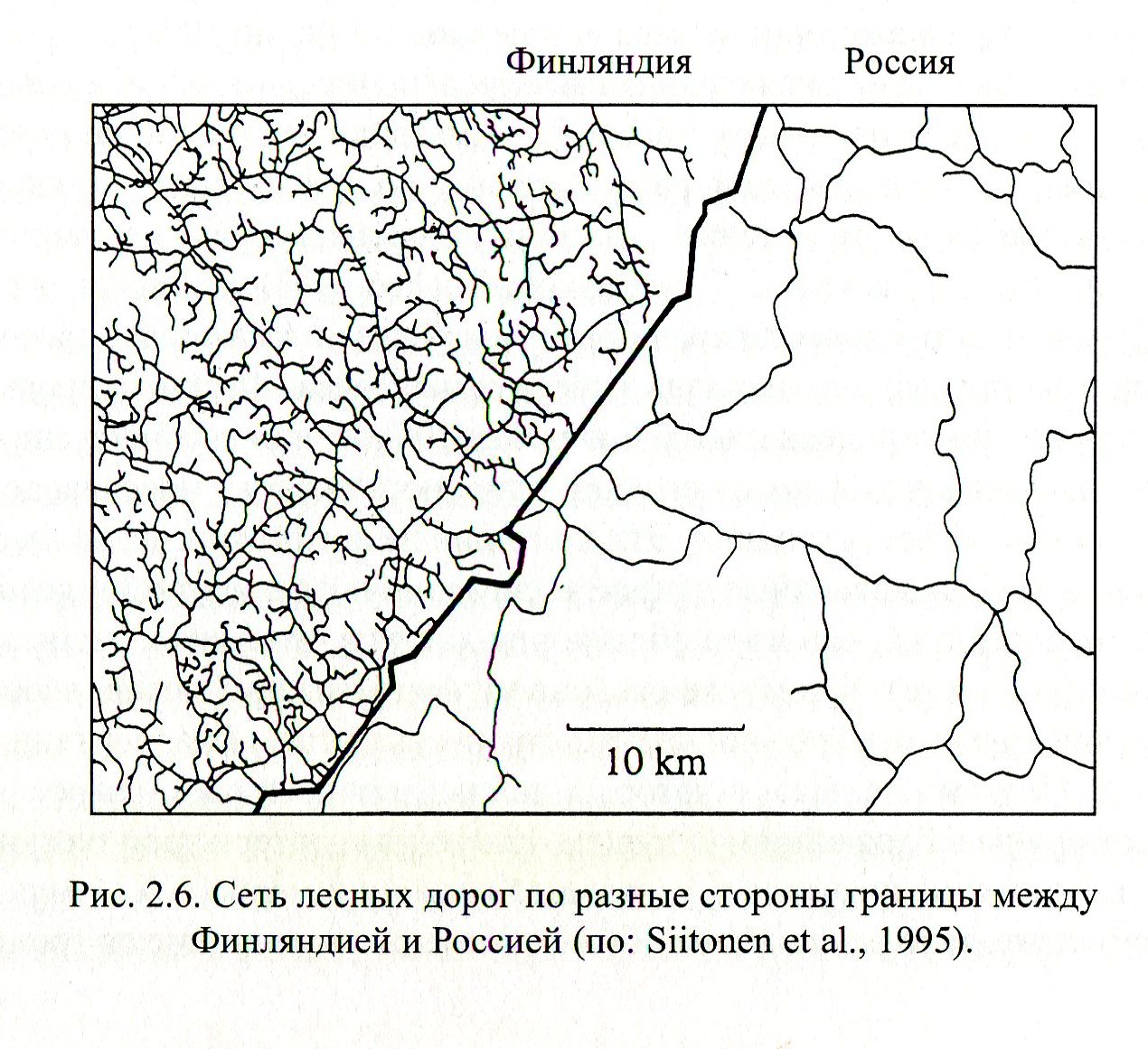
Источник. Илкка Хански. Ускользающий мир. Экологические последствия утраты местообитаний. М.: Товарищество научных изданий, 2010. С.109..
В продолжение темы «Капитализм против природы»
Я писал, что Финляндия - отличный пример, как в условиях рыночной экономики при слабом противодействии природоохранников можно изгадить почти девственную природу за очень короткий период промышленного развития. Достаточно сравнить степень фрагментации местообитаний дорожной сетью (которая определяет и уязвимость видов к воздействию человека, и риск их выпадения из состава фауны по случайным причинам, согласно моделям островной биогеографии) по обе стороны советско-финской границы.
Хотя урбанизация и промышленное развитие Русской Карелии сравнимо с таковым Финской, а то и превышает его, фрагментированность естественных местообитаний во втором случае на порядок больше. См.например, постоянно растущую с 1945 по 1995 год фрагментацию старовозрастного елового леса в Кухмо (восточная Финляндия).

Источник. Илкка Хански, 2010, ibid.
Притом что индустриализация Русской Карелии началась раньше, и «нагруженность» индустрией городских центров там была выше, малонарушенная природная периферия региона (а, следовательно, и биоразнообразие) сохраняется лучше.
История природопользования в Финляндии интересна ещё в одном отношении. Благодаря хорошим рабочим контактам наших охотоведов с финскими в 1970-е гг. Финляндия - одна из немногих стран Запада, использующая наш метод оценки обилия дичи по зимним маршрутным учётам (ЗМУ[1]).
Это единственный в настоящее время метод массового учёта охотничьих животных, позволяющий давать оценки хотя бы относительного обилия (выражается в числе следов на 10 км маршрута за сутки) для довольно широкого спектра активных в зимнее время видов. Соответственно, есть возможность сравнить интенсивности вырубки леса в наших странах, связанной с развитием приграничных территорий, и степень экологичности рубок, по степени влияния на популяции боровой дичи и охотничьих зверей.
Это сравнение проведено в работе Ю.П.Курхинена с соавт.[2] 1999 г.; ими впервые предлагается по данным ЗМУ провести анализ видового богатства охотничьей фауны по признаку «наличия-отсутствия» в квадрате того или иного вида за 5 лет (1990-1994 гг.): а) из общего списка в 20 видов (группа «всех» видов, включая 4 вида лесных тетеревиных птиц); б) из списка в 8 таёжных видов - белка, куница, росомаха, волк, бурый медведь, рысь, дикий лесной северный олень, глухарь. Таким образом, поквадратно оценивались индексы видового богатства для группы «всех» видов и отдельно - для условно названной группы «лесных» видов, а также индексы относительного обилия по ЗМУ в среднем за 5 лет. Общий объём зимних учётов в районе исследований (Карелия и Восточная Финляндия) за 1990-1994 гг. составил 77876 км.
Авторы использовали показатель «лесоэксплуатационной нагрузки на экосистемы«, выражающего объём древесины, изымаемой с 1 га лесной площади. Динамика роста этого показателя в Финляндии и в Российской Карелии была сходной и связанной с ростом промышленного производства (коэффициент корреляции за период 1955-1992 гг. составил +0.54; P<0,001). Однако интенсивность лесоэксплуатационных нагрузок в Карелии была в 1.6 раза ниже, 144 м3/га и 235 м3/га соответственно. И самая главное, она имела более благоприятный для фауны и лесовозобновления пространственный паттерн. Невырубленные участки оставались связными, не образовывали «архипелагов», тем самым не «подставляли» первое и второе под негативное воздействие островного эффекта[3].
В сравнении с Финляндией освоение лесов в в Карели и было более неоднородным как в территориальном, так и временном аспектах: интенсивные рубки леса распространялись «волнами» от крупных транспортных узлов вглубь территории, с сохранением малонарушенной природной периферии, поддерживающей связность популяций диких видов. В Финляндии освоение было сплошным, а крайне развивая дорожная сеть рассекала периферию каждого региона (поскольку была развита там не хуже, чем вблизи центров), не позволяя ей функционировать как канал связи между популяциями сохранившихся лесных участков. Также коэффициент вариации лесоэксплуатационных нагрузок по годам для Карелии был в 2.2 раза выше, чем в Финляндии. Благодаря этому популяции охотничьих видов могли восстанавливаться между пиковыми годами, при одновременном снижении посещаемости лесов лесорубами и промысловиками, существенно понижающем риск пожаров.
«Очень важны различия в технологии лесозаготовок и восстановления леса. В лесах Карелии 3й группы, включающих более 80% лесного фонда, разрешались практически неограниченные по площади сплошные концентрированные рубки, которые осуществлялись крупными механизированными бригадами на тяжёлых гусеничных тракторах. Массовыми являлись оставление недорубов и сохранение семенных куртин. Обязательным был сбор и сжигание порубочных остатков. Восстановление леса происходило естественным путём, рубки ухода не имели существенного значен7ия в динамике состава лесов. В северной части региона болота слабо затрагивались лесоосушительной мелиорацией, дорожная сеть была развита слабо и после освоения массивов постепенно разрушалась». Иными словами, массивы не затронутые рубками, как и восстановившиеся после рубок, сближались с малонарушенными лесами по таким важным параметрам, как обилие мёртвой древесины, разномасштабная мозаичность, в том числе образованная «вкраплениями» лист венных пород, представленность болотных массивов.
«В Финляндии практиковались менее крупные вырубки, измельчение или сжигание порубочных остатков, преимущественно искусственное лесовозобновление. Здесь существовала сеть постоянно действующих дорог, проводились интенсивные рубки ухода и повсеместная лесоосушительная мелиорация. Так, к началу 1990-х гг. здесь осушено и переведено в категорию лесов 6 из 10 млн.га болотных земель».
Соответственно, эти леса максимально по биоценотически важным параметрам максимально приближены к коммерческим[4], проблемным с точки зрения сохранения биоразнообразия. Во-вторых, фрагментация на низовом уровне в них крайне велика, что делает фауну данных местообитаний исключительно уязвимой к островному эффекту, одновременно препятствуя сохранению крупных массивов старых лесов и верховых болот. При рубках в Карелии они сохраняются, благодаря чему фауна, в том числе охотничья, «отступая» туда, может выждать восстановление лесов после рубок, чем снижается риск полного исчезновения видов на соответствующей территории. Кроме того, нерубленные участки вместе с восстановившимися в Карелии сохраняют связность по периферии, чем образуются «таёжные коридоры[5]», крайне важные для сохранения фауны в условиях фрагментирующего воздействия рубок.
Благодаря этому при лесоэксплуатационных нагрузках, различающихся всего в 1.6 раза, лесные ландшафты Финляндии показывают качественно иной (много больший) уровень антропогенной трансформации. В сравнении с Карелией, в них существенно ниже представленность важных для фауны ключевых местообитаний - спелых лесов (старше 100 лет), ельников и особенно сосняков, болот и «неудобий» (участков, вообще не нарушенных лесоэксплуатацией). В Финляндии, соответственно, существенно больше сельхозземель на месте осушенных болот, дорог и населённых пунктов, а также территорий, занятых коммерческими лесами. Особенно важно различие в представленности спелых сосняков - местообитания, наиболее важного для охотничьей фауны, а также болот. Они хорошо сохранились в Карелии и плохо - в Финляндии, где основной удар лесоэксплуатационного освоения территории пришёлся на эти местообитания (в Карелии - на все равномерно, нет такой избирательности).
«В результате существенных различий в формах, интенсивности и технологии лесопользования, в Финляндии и на северо-западе России сформировались ландшафты разные по структуре местообитаний таёжных животных. В Финляндии это преимущественно мозаичные, мелкоконтурные леса с небольшими вырубками, широким спектром относительно равномерно представленных древостоев на разных стадиях сукцессии, небольшой долей лесов в возрасте более 100 лет, хорошей транспортной и сельскохозяйственной освоенностью территории. В Российской Карелии это чаще всего неоднородный ландшафт, с наличием как крупных вырубок, так и значительных массивов спелых лесов, в том числе в виде недорубов, семенных куртин, водоохранных полос и заболоченных древостоев, сравнительно плохим состоянием лесовозных дорог, слабой сельскохозяйственной освоенностью территории. Так, доля лесов старше 100 лет и болот в Карелии соответственно в 1.6 и 5.2 раза выше, тогда как в Восточной Финляндии в 2.5, 3.2 и 28 (!) раз выше представленность застроенных земель, дорог и сельхозугодий. Превышение (в 1.4 раза) доли средневозрастных древостоев (40-100 лет) в Финляндии по сравнению с Карелией фактически отражает различия в интенсивности лесопользования в течение 20 века. Более высокая (в 1.5 раза) доля лиственных лесов в Российской Карелии при несколько более низкой доле сосняков хорошо иллюстрирует разную стратегию лесовосстановления в этих регионах в послевоенные годы. Это фактически повсеместное естественное восстановление со сменой пород в Карелии и массовое создание лесных культур сосны в Восточной Финляндии».
Однако доля спелых лесов в Карелии выше - и особенно сосновых боров, крайне важных для дичи. «В Восточной Финляндии суммарная доля спелых лесов, болот и «неудобий» (т.н. «ненарушенные территории) в 3 раза уступает антропогенным территориям (вырубки, дороги и пр.). В Карелии это соотношение близко к 1:1. Приверженность к сравнительно эффективной «скандинавской» технологии лесопользования в Финляндии более негативно отразилось на состоянии местообитаний и разнообразии охотничьей фауны, чем «экстенсивные» формы лесоэксплутатации, практиковавшиеся в Российской Карелии Советского периода.
Так, общее видовое богатство охотничьих животных позитивно связано с долей спелых хвойных лесов, а индекс видового богатства группы «лесных» видов имеет сильную обратную зависимость от интенсивности лесохозяйственного освоения территории, выраженной в представительности вторичных средневозрастных древостоев. Ещё более важные взаимосвязи проявляются, если рассматривать региональную специфику влияния участков спелых лесов с учётом их породного состава. Оказывается, что наиболее сильное позитивное влияние на индекс видового богатства животных оказывает представленность сосняков возрастной категории старше 100 лет. Для спелых лиственных лесов и для ельников данной возрастной категории его вообще практически нет. Последнее отчасти определяется тем, что наиболее требовательные к почвам ельники и лиственные леса расположены в тех регионах, которые историче5ски ранее и активнее других осваивались человеком (с соответствующими н7егативными последствиями для таёжной фауны). Это подтверждается наличием прямой связи представленности спелых ельников и лиственных лесов с уровнем антропогенной трансформированности территории - коэффициенты корреляции, соответственно, +0.57 и +0.61; Р<0,01. Для сосняков эта связь однозначно обратная (-0.56).
Таким образом, районы с наибольшим разнообразием охотничьих животных совпадают с наибольшей представленностью спелых сосняков.
В этом случае не акцентируется обязательность наличия в прямых регрессионных зависимостях только одного фактора (спелые леса), поскольку здесь должен играть определяющую роль весь комплекс показателей, характеризующих структуру ландшафта. Так, в «лучших» квадратах (с максимальным индексом богатства охотничьих видов) по сравнению с «худшими» была значительно выше представленность не только спелых сосняков (в 2.5 раза), но и также болот (в 6 раз), и более чем в 20 (!) раз ниже сельскохозяйственная освоенность территории.
Коренные леса - в данном случае один из ключевых признаков, характеризующих степень сохранности структуры аборигенного ландшафта. Разнонаправленность связи (смена знака с преимущественно «плюс» в Восточной Финляндии на «минус» в Российской Карелии) - один из важных признаков региональной специфики указанных здесь взаимосвязей, который имеет свой экологический смысл…
Вполне очевидно, что между этими двумя разнонаправленными зависимостями должна существовать «критическая зона» перехода из условно оптимальной (Средняя Карелия) в условно пессимальную зону. Более подробный регрессионный анализ показывает, что фактически существует 3 зоны, характеризующихся различной представленностью хвойных лесов старших возрастных групп и, что особенно важно, разным характером взаимосвязей между долей спелых лесов и видовым разнообразием животных.
В первой зоне «относительного благополучия» эта доля свыше 80% и здесь не отмечено устойчивых корреляционных связей между данным показателем и видовым разнообразием. Во второй «переходной» зоне доля таких лесов 10-20% и это приводит к некоторому увеличению видового богатства (коэффициент корреляции - -0.43). Авторы показывают, что это происходит за счёт «псевдообогащения» фауны, скажем, за счёт иммиграции некоторых южных видов, но число специализированных таёжных видов сокращается.
Трансформация коренных лесов в «критической» зоне (<10%) ведёт к прогрессирующему сокращению видового богатства и разнообразия (коэффициент корреляции - +0.5). Это характерно для наиболее освоенных территорий финляндской части региона».
Иными словами, финское лесное хозяйство, управляемое рыночной экономикой, в сравнении с советским плановым, не только истощительней само по себе, но и «тянет» за собой сельскохозяйственную трансформацию лесного ландшафта, развитие дорожной сети и пр. трансформации. Они дают возможность эксплуатировать данную территорию другими способами, почему она теряет качество местообитания для ценных видов даже в случае, когда лес формально восстанавливается. Напротив, советский тип эксплуатации сохраняет лесной ландшафт не только как лес, но и как местообитания охотничьих видов (и прочих видов, составляющих исходное биоразнообразие данной территории), то есть существенно безопасней в экологическом плане.
+интересная дискуссия в комментах
Примечания:
[1] см. также «Методические рекомендации по организации, проведению и обработке результатов ЗМУ охотничьих животных в РФ» 2009 г, и основную работу С.Г.Приклонского, 1973. Зимний маршрутный учет охотничьих животных // Тр. Окскогого зап. Вып. 9. С. 35-62.
[2] Курхинен Ю.П., Волков А.Д, Данилов П.И., Громцев А.Н., Линден Х., Хелле П. Значение коренных лесов для сохранения видового разнообразия охотничьих животных в Карелии и Восточной Фенноскандии // Коренные леса таежной зоны Европы: современное состояние и проблемы сохранения. Петрозаводск, 1999. C. 27-31.
[3] См. « Большой минус, маленький плюс - экологические последствия фрагментации местообитаний»
[4] См. « Чем коммерческий лес отличается от ненарушенного?»
[5] См. Ю.Курхинен, А.Н.Громцев, П.И.Данилов, А.М.Крышень, Х.Линден, Т.Линдхольм, 2009. «Особенности и значение таёжных коридоров в Восточной Фенноскандии»// Труды Карелского научного центра РАН. №2. С.16-23.
http://www.socialcompas.com/2014/09/24/e-kosistemny-e-posledstviya-lesoe-kspluatatsii-vostochnaya-finlyandiya-vs-kareliya/
Источник. Илкка Хански. Ускользающий мир. Экологические последствия утраты местообитаний. М.: Товарищество научных изданий, 2010. С.109..
В продолжение темы «Капитализм против природы»
Я писал, что Финляндия - отличный пример, как в условиях рыночной экономики при слабом противодействии природоохранников можно изгадить почти девственную природу за очень короткий период промышленного развития. Достаточно сравнить степень фрагментации местообитаний дорожной сетью (которая определяет и уязвимость видов к воздействию человека, и риск их выпадения из состава фауны по случайным причинам, согласно моделям островной биогеографии) по обе стороны советско-финской границы.
Хотя урбанизация и промышленное развитие Русской Карелии сравнимо с таковым Финской, а то и превышает его, фрагментированность естественных местообитаний во втором случае на порядок больше. См.например, постоянно растущую с 1945 по 1995 год фрагментацию старовозрастного елового леса в Кухмо (восточная Финляндия).
Источник. Илкка Хански, 2010, ibid.
Притом что индустриализация Русской Карелии началась раньше, и «нагруженность» индустрией городских центров там была выше, малонарушенная природная периферия региона (а, следовательно, и биоразнообразие) сохраняется лучше.
История природопользования в Финляндии интересна ещё в одном отношении. Благодаря хорошим рабочим контактам наших охотоведов с финскими в 1970-е гг. Финляндия - одна из немногих стран Запада, использующая наш метод оценки обилия дичи по зимним маршрутным учётам (ЗМУ[1]).
Это единственный в настоящее время метод массового учёта охотничьих животных, позволяющий давать оценки хотя бы относительного обилия (выражается в числе следов на 10 км маршрута за сутки) для довольно широкого спектра активных в зимнее время видов. Соответственно, есть возможность сравнить интенсивности вырубки леса в наших странах, связанной с развитием приграничных территорий, и степень экологичности рубок, по степени влияния на популяции боровой дичи и охотничьих зверей.
Это сравнение проведено в работе Ю.П.Курхинена с соавт.[2] 1999 г.; ими впервые предлагается по данным ЗМУ провести анализ видового богатства охотничьей фауны по признаку «наличия-отсутствия» в квадрате того или иного вида за 5 лет (1990-1994 гг.): а) из общего списка в 20 видов (группа «всех» видов, включая 4 вида лесных тетеревиных птиц); б) из списка в 8 таёжных видов - белка, куница, росомаха, волк, бурый медведь, рысь, дикий лесной северный олень, глухарь. Таким образом, поквадратно оценивались индексы видового богатства для группы «всех» видов и отдельно - для условно названной группы «лесных» видов, а также индексы относительного обилия по ЗМУ в среднем за 5 лет. Общий объём зимних учётов в районе исследований (Карелия и Восточная Финляндия) за 1990-1994 гг. составил 77876 км.
Авторы использовали показатель «лесоэксплуатационной нагрузки на экосистемы«, выражающего объём древесины, изымаемой с 1 га лесной площади. Динамика роста этого показателя в Финляндии и в Российской Карелии была сходной и связанной с ростом промышленного производства (коэффициент корреляции за период 1955-1992 гг. составил +0.54; P<0,001). Однако интенсивность лесоэксплуатационных нагрузок в Карелии была в 1.6 раза ниже, 144 м3/га и 235 м3/га соответственно. И самая главное, она имела более благоприятный для фауны и лесовозобновления пространственный паттерн. Невырубленные участки оставались связными, не образовывали «архипелагов», тем самым не «подставляли» первое и второе под негативное воздействие островного эффекта[3].
В сравнении с Финляндией освоение лесов в в Карели и было более неоднородным как в территориальном, так и временном аспектах: интенсивные рубки леса распространялись «волнами» от крупных транспортных узлов вглубь территории, с сохранением малонарушенной природной периферии, поддерживающей связность популяций диких видов. В Финляндии освоение было сплошным, а крайне развивая дорожная сеть рассекала периферию каждого региона (поскольку была развита там не хуже, чем вблизи центров), не позволяя ей функционировать как канал связи между популяциями сохранившихся лесных участков. Также коэффициент вариации лесоэксплуатационных нагрузок по годам для Карелии был в 2.2 раза выше, чем в Финляндии. Благодаря этому популяции охотничьих видов могли восстанавливаться между пиковыми годами, при одновременном снижении посещаемости лесов лесорубами и промысловиками, существенно понижающем риск пожаров.
«Очень важны различия в технологии лесозаготовок и восстановления леса. В лесах Карелии 3й группы, включающих более 80% лесного фонда, разрешались практически неограниченные по площади сплошные концентрированные рубки, которые осуществлялись крупными механизированными бригадами на тяжёлых гусеничных тракторах. Массовыми являлись оставление недорубов и сохранение семенных куртин. Обязательным был сбор и сжигание порубочных остатков. Восстановление леса происходило естественным путём, рубки ухода не имели существенного значен7ия в динамике состава лесов. В северной части региона болота слабо затрагивались лесоосушительной мелиорацией, дорожная сеть была развита слабо и после освоения массивов постепенно разрушалась». Иными словами, массивы не затронутые рубками, как и восстановившиеся после рубок, сближались с малонарушенными лесами по таким важным параметрам, как обилие мёртвой древесины, разномасштабная мозаичность, в том числе образованная «вкраплениями» лист венных пород, представленность болотных массивов.
«В Финляндии практиковались менее крупные вырубки, измельчение или сжигание порубочных остатков, преимущественно искусственное лесовозобновление. Здесь существовала сеть постоянно действующих дорог, проводились интенсивные рубки ухода и повсеместная лесоосушительная мелиорация. Так, к началу 1990-х гг. здесь осушено и переведено в категорию лесов 6 из 10 млн.га болотных земель».
Соответственно, эти леса максимально по биоценотически важным параметрам максимально приближены к коммерческим[4], проблемным с точки зрения сохранения биоразнообразия. Во-вторых, фрагментация на низовом уровне в них крайне велика, что делает фауну данных местообитаний исключительно уязвимой к островному эффекту, одновременно препятствуя сохранению крупных массивов старых лесов и верховых болот. При рубках в Карелии они сохраняются, благодаря чему фауна, в том числе охотничья, «отступая» туда, может выждать восстановление лесов после рубок, чем снижается риск полного исчезновения видов на соответствующей территории. Кроме того, нерубленные участки вместе с восстановившимися в Карелии сохраняют связность по периферии, чем образуются «таёжные коридоры[5]», крайне важные для сохранения фауны в условиях фрагментирующего воздействия рубок.
Благодаря этому при лесоэксплуатационных нагрузках, различающихся всего в 1.6 раза, лесные ландшафты Финляндии показывают качественно иной (много больший) уровень антропогенной трансформации. В сравнении с Карелией, в них существенно ниже представленность важных для фауны ключевых местообитаний - спелых лесов (старше 100 лет), ельников и особенно сосняков, болот и «неудобий» (участков, вообще не нарушенных лесоэксплуатацией). В Финляндии, соответственно, существенно больше сельхозземель на месте осушенных болот, дорог и населённых пунктов, а также территорий, занятых коммерческими лесами. Особенно важно различие в представленности спелых сосняков - местообитания, наиболее важного для охотничьей фауны, а также болот. Они хорошо сохранились в Карелии и плохо - в Финляндии, где основной удар лесоэксплуатационного освоения территории пришёлся на эти местообитания (в Карелии - на все равномерно, нет такой избирательности).
«В результате существенных различий в формах, интенсивности и технологии лесопользования, в Финляндии и на северо-западе России сформировались ландшафты разные по структуре местообитаний таёжных животных. В Финляндии это преимущественно мозаичные, мелкоконтурные леса с небольшими вырубками, широким спектром относительно равномерно представленных древостоев на разных стадиях сукцессии, небольшой долей лесов в возрасте более 100 лет, хорошей транспортной и сельскохозяйственной освоенностью территории. В Российской Карелии это чаще всего неоднородный ландшафт, с наличием как крупных вырубок, так и значительных массивов спелых лесов, в том числе в виде недорубов, семенных куртин, водоохранных полос и заболоченных древостоев, сравнительно плохим состоянием лесовозных дорог, слабой сельскохозяйственной освоенностью территории. Так, доля лесов старше 100 лет и болот в Карелии соответственно в 1.6 и 5.2 раза выше, тогда как в Восточной Финляндии в 2.5, 3.2 и 28 (!) раз выше представленность застроенных земель, дорог и сельхозугодий. Превышение (в 1.4 раза) доли средневозрастных древостоев (40-100 лет) в Финляндии по сравнению с Карелией фактически отражает различия в интенсивности лесопользования в течение 20 века. Более высокая (в 1.5 раза) доля лиственных лесов в Российской Карелии при несколько более низкой доле сосняков хорошо иллюстрирует разную стратегию лесовосстановления в этих регионах в послевоенные годы. Это фактически повсеместное естественное восстановление со сменой пород в Карелии и массовое создание лесных культур сосны в Восточной Финляндии».
Однако доля спелых лесов в Карелии выше - и особенно сосновых боров, крайне важных для дичи. «В Восточной Финляндии суммарная доля спелых лесов, болот и «неудобий» (т.н. «ненарушенные территории) в 3 раза уступает антропогенным территориям (вырубки, дороги и пр.). В Карелии это соотношение близко к 1:1. Приверженность к сравнительно эффективной «скандинавской» технологии лесопользования в Финляндии более негативно отразилось на состоянии местообитаний и разнообразии охотничьей фауны, чем «экстенсивные» формы лесоэксплутатации, практиковавшиеся в Российской Карелии Советского периода.
Так, общее видовое богатство охотничьих животных позитивно связано с долей спелых хвойных лесов, а индекс видового богатства группы «лесных» видов имеет сильную обратную зависимость от интенсивности лесохозяйственного освоения территории, выраженной в представительности вторичных средневозрастных древостоев. Ещё более важные взаимосвязи проявляются, если рассматривать региональную специфику влияния участков спелых лесов с учётом их породного состава. Оказывается, что наиболее сильное позитивное влияние на индекс видового богатства животных оказывает представленность сосняков возрастной категории старше 100 лет. Для спелых лиственных лесов и для ельников данной возрастной категории его вообще практически нет. Последнее отчасти определяется тем, что наиболее требовательные к почвам ельники и лиственные леса расположены в тех регионах, которые историче5ски ранее и активнее других осваивались человеком (с соответствующими н7егативными последствиями для таёжной фауны). Это подтверждается наличием прямой связи представленности спелых ельников и лиственных лесов с уровнем антропогенной трансформированности территории - коэффициенты корреляции, соответственно, +0.57 и +0.61; Р<0,01. Для сосняков эта связь однозначно обратная (-0.56).
Таким образом, районы с наибольшим разнообразием охотничьих животных совпадают с наибольшей представленностью спелых сосняков.
В этом случае не акцентируется обязательность наличия в прямых регрессионных зависимостях только одного фактора (спелые леса), поскольку здесь должен играть определяющую роль весь комплекс показателей, характеризующих структуру ландшафта. Так, в «лучших» квадратах (с максимальным индексом богатства охотничьих видов) по сравнению с «худшими» была значительно выше представленность не только спелых сосняков (в 2.5 раза), но и также болот (в 6 раз), и более чем в 20 (!) раз ниже сельскохозяйственная освоенность территории.
Коренные леса - в данном случае один из ключевых признаков, характеризующих степень сохранности структуры аборигенного ландшафта. Разнонаправленность связи (смена знака с преимущественно «плюс» в Восточной Финляндии на «минус» в Российской Карелии) - один из важных признаков региональной специфики указанных здесь взаимосвязей, который имеет свой экологический смысл…
Вполне очевидно, что между этими двумя разнонаправленными зависимостями должна существовать «критическая зона» перехода из условно оптимальной (Средняя Карелия) в условно пессимальную зону. Более подробный регрессионный анализ показывает, что фактически существует 3 зоны, характеризующихся различной представленностью хвойных лесов старших возрастных групп и, что особенно важно, разным характером взаимосвязей между долей спелых лесов и видовым разнообразием животных.
В первой зоне «относительного благополучия» эта доля свыше 80% и здесь не отмечено устойчивых корреляционных связей между данным показателем и видовым разнообразием. Во второй «переходной» зоне доля таких лесов 10-20% и это приводит к некоторому увеличению видового богатства (коэффициент корреляции - -0.43). Авторы показывают, что это происходит за счёт «псевдообогащения» фауны, скажем, за счёт иммиграции некоторых южных видов, но число специализированных таёжных видов сокращается.
Трансформация коренных лесов в «критической» зоне (<10%) ведёт к прогрессирующему сокращению видового богатства и разнообразия (коэффициент корреляции - +0.5). Это характерно для наиболее освоенных территорий финляндской части региона».
Иными словами, финское лесное хозяйство, управляемое рыночной экономикой, в сравнении с советским плановым, не только истощительней само по себе, но и «тянет» за собой сельскохозяйственную трансформацию лесного ландшафта, развитие дорожной сети и пр. трансформации. Они дают возможность эксплуатировать данную территорию другими способами, почему она теряет качество местообитания для ценных видов даже в случае, когда лес формально восстанавливается. Напротив, советский тип эксплуатации сохраняет лесной ландшафт не только как лес, но и как местообитания охотничьих видов (и прочих видов, составляющих исходное биоразнообразие данной территории), то есть существенно безопасней в экологическом плане.
+интересная дискуссия в комментах
Примечания:
[1] см. также «Методические рекомендации по организации, проведению и обработке результатов ЗМУ охотничьих животных в РФ» 2009 г, и основную работу С.Г.Приклонского, 1973. Зимний маршрутный учет охотничьих животных // Тр. Окскогого зап. Вып. 9. С. 35-62.
[2] Курхинен Ю.П., Волков А.Д, Данилов П.И., Громцев А.Н., Линден Х., Хелле П. Значение коренных лесов для сохранения видового разнообразия охотничьих животных в Карелии и Восточной Фенноскандии // Коренные леса таежной зоны Европы: современное состояние и проблемы сохранения. Петрозаводск, 1999. C. 27-31.
[3] См. « Большой минус, маленький плюс - экологические последствия фрагментации местообитаний»
[4] См. « Чем коммерческий лес отличается от ненарушенного?»
[5] См. Ю.Курхинен, А.Н.Громцев, П.И.Данилов, А.М.Крышень, Х.Линден, Т.Линдхольм, 2009. «Особенности и значение таёжных коридоров в Восточной Фенноскандии»// Труды Карелского научного центра РАН. №2. С.16-23.
http://www.socialcompas.com/2014/09/24/e-kosistemny-e-posledstviya-lesoe-kspluatatsii-vostochnaya-finlyandiya-vs-kareliya/