Лепоспондилы. Скелет - дело тонкое!
Подкласс лепоспондил - палеозойские амфибии, которые, в противоположность темноспондилам и рептилиоморфам, не упрочняли и наращивали скелет, а облегчали. Отсюда и название - тонкопозвонковые. Позвонки у них - тонкая катушка вокруг хорды и спинного мозга. Зато их много (у некоторых более двухсот). А это существенно повышает гибкость в теле.

Чешские лепоспондилы позднего карбона от Зденека Буриана. Справа на камешке темноспондил Branchiosaurus salamandroides, слева - Phlegethontia longissimum (Aistopoda), вверху - Sauropleura scalaris (Nectridea), внизу - Microbranchius pelicani (Microsauria)
Облегчение позвоночника означает, что мы больше не удерживаем тело на весу - а значит, не ходим, а ползаем, роем или плаваем. Лапы нам при таком подходе не особо нужны - и они редуцируются, вплоть до полного исчезновения. У плавающих форм могут сохраняться развитые задние лапки, у роющих - передние.
Вес тела при этом должен быть невелик - и самый крупный лепоспондил достигает длины лишь в метр. Зато очень маленьких существ может быть ОЧЕНЬ много. Поэтому, несмотря на плохую сохраняемость мелких тонких косточек, останки лепоспондил в заметных количествах находят в Северной Америке, Северной Африке и Европе.
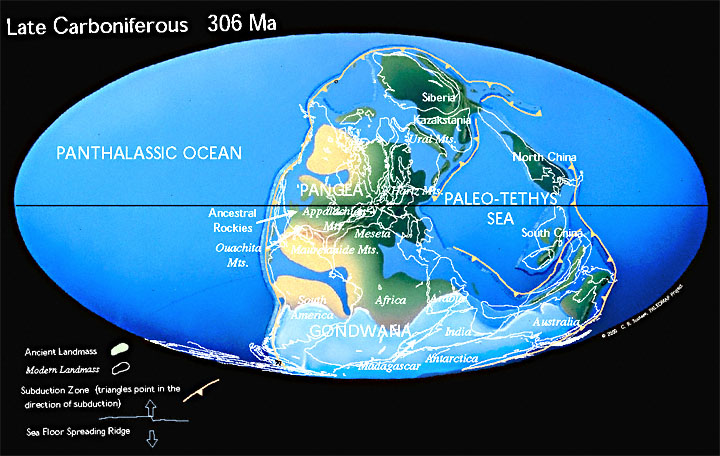
Поздний карбон, образование Пангеи и оледенение. Европа и Северная Америка - в центре
Филогенетическое положение лепоспондил по сей день остается неясным. Из полусотни крупных таксонов позвоночных только они и анапсиды (к которым относятся черепахи) - темна вода в облацех.
Во-первых, нам совершенно неизвестны предковые виды лепоспондил - они обнаруживаются в карбоне as is уже готовыми, специализированными. А в девоне мы имеем только переходные формы, общие для всех четвероногих.
Во-вторых, отследить их потомков и братьев непросто, поскольку любое сходство может быть конвергентным. То есть, признаки совпадают не потому, что позднейшее существо происходит от примитивной амфибии, а просто в силу сходного образа жизни и решения одних и тех же задач.
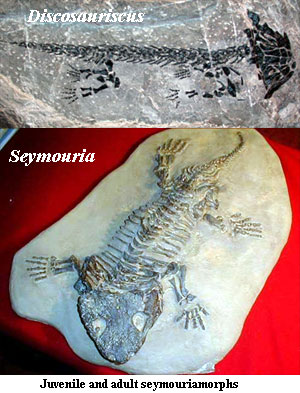
Ювенильная и взрослая формы сеймурии
В-третьих, земноводные в своем индивидуальном развитии принимают две совершенно разные формы - водную ювенильную ("головастик") и половозрелую. И лепоспондилы, упростившись, педоморфны (сохраняют признаки детской формы). Не зная настоящей взрослой формы животного, как делать выводы об их систематическом положении?

Поролепиформные рыбы
Исследование кистеперых рыб показало, что тонкопозвонковые напоминают поролепиформных рыб, а не остеолепиоморфных, от которых произошли тетраподы. Получается, что лепоспондилы могут быть совершенно отдельной ветвью эволюции наземных позвоночных. Однако, оппоненты говорят, что тонкий позвоночник вполне мог возникать независимо вследствие педоморфоза. Тогда лепоспондилы могут оказаться полифилетической (не имеющей единого предка) группой. Общепринятая точка зрения - что они все-таки близкие родственники рептилиоморф и имеют одного общего предка.

Угольные леса.
Ранее некоторые считали лепоспондил предками только саламандр. С объединением лягушек, саламандр и червяг в подкласс беспанцирных от этой точки зрения отказались. Лепоспондил стали считать предками всех современных амфибий. Принятая сейчас "темноспондильная" гипотеза отказывает тонкопозвонковым в наследниках, полагая их тупиковой ветвью. Впрочем, появление доказательств отдаленности червяг от общего тренда развития голых земноводных придало убедительности гипотезе, что они таки происходят от лепоспондил, а вот все остальные - нет.

Филогенетическое древо лепоспондильной гипотезы
Традиционно базальной формой лепоспондил считались микрозавры - "крошечные ящерицы", с коротким туловищем и маленькими лапками. Добрый дедушка Кэрролл (написавший трехтомник о эволюции позвоночных) разделил их на два подотряда - тудитаноморф и микробрахоморф. Различить подотряды трудно, да, в общем, уже и не нужно. Современные кладисты очень любят менять положение групп туда-сюда и пуристически уничтожать их за грех парафилии, но вот написать толковое описание - этого от них хрен дождешься. Поэтому приходится пользоваться древней классификацией (вот тут лежит гроссбух Кэрролла и Гаскелл по микрозаврам на аглицком)
Подотряд микробрахоморф - мелкие и очень мелкие позвоночные с костяными пластинами на коже, напоминающими чешую рептилий. Микробрахоморфы практически упразднены современной систематикой, поскольку в их состав входят базальные формы микрозавров и довольно специализированные, далеко отстоящие от них брахистелехиды. Череп треугольный, шире чем у тудитаноморф, отверстия лишь для ноздрей и больших глазниц. Внутри черепа хорошо развитая "ушная капсула" - возможно здесь находилось внутреннее ухо. Морда короткая, глаза сбоку. Малое количество простых конических зубов, несколько могут быть увеличенными (напоминающими клыки), напоминает современных земноводных. Позвоночник гибкий и слабый, у пермских видов становится короче и прочнее.

Ютагерпетон (318-314 миллионов лет, Юта) - базальная форма лепоспондил, отдельные товарищи даже не причисляют их поэтому к микрозаврам

Гилоплезион (311-307 миллионов лет, Чехия) - задние лапы длиннее передних, насчитывающих лишь три пальца. Образ жизни неясен, мог быть как водным, так и наземным

Одонтерпетон (311-307 млн. лет, Огайо) - найдена только передняя половина скелета. Предположительно продолговатое тело 15 см длиной, три или четыре пальца на передних лапах.

Микробрахис (311-307 миллионов лет, Чехия) - саламандроподобное существо 15 сантиметров длиной, по видимому плавал как рыба, латеральными движениями тела и питался планктоном.
В перми семейство брахистелехид с тремя родами - батропеты (4 вида, Германия), монотипичные кэрролла (Техас, 290-279 млн лет) и квазицецилия (псевдочервяга, Техас, 279-272 млн лет)

Батропет (299-290 млн лет, Германия) - примерно 8 см длиной, череп короткий с большими орбитами.
Подотряд тудитаноморфы объединяет 7 семейств (из них два - монотипические). Вероятно у них не было водной личиночной стадии, либо она была краткой. Если верно первое, то их нельзя считать амфибиями и это параллельная поытка "амниотизации" - полностью наземной формы позвоночных. Лепоспондильный позвоночник не мог нести вес тела и спинные отростки разрастались продольно, налегая друг на друга и принимая нагрузку на себя. Хвост не приспособлен для плаванья, на нем нет нижних отростков. Конечности короткие, у большинства расположены по бокам в нижней части туловища, позволяли опираться на них и передвигаться по суше. Кожа многих несла костяные чешуи, обычно они формировали костяной панцирь с брюшной стороны, чтобы защитить ее от повреждений, но иногда были и на спине.
Форма черепа треугольная, морда короткая, заглазничная часть длиннее морды. Крупные, круглые глазницы ориентированы вперед и в стороны, что дает бинокулярное зрение в переднем направлении. Окон в черепе для выхода жевательных мышц не было, поэтому он сильно расширен в щечной области, чтобы поместить всю жевательную мускулатуру под крышей. У некоторых более поздних форм в боковой поверхности появилась щечная врезка. Зубы простые конические, у некоторых мелкие, у других - крупные, с более тупыми вершинами. Имелись небные зубы, иногда часто рассеяные по небу, иногда формирующие параллельный ряд зубов в верхней челюсти. Все это позволяло пережевывать хитин насекомых.
Семейство Гапсидопарейонтид (318-272 млн лет, Оклахома, Чехия, Германия, Канада) имело большую щечную врезку, где могла размещаться сильная жевательная мускулатура

Ллистроф (290-268 млн лет, Оклахома) - самый крупный представитель семейства, не более 20 см. Ноги короткие, но хорошо сформированные

Саксонерпетон (299-295 млн. лет, Саксония) - отсутствие щечной врезки и возможное наличие органа боковой линии намекает на то, что либо это базальная форма, либо вообще не гапсидопарейонтид.
Семейство тудитан (311-303 млн. лет, Огайо, Чехия, Канада) напоминало ящериц среднего размера. И не только внешне. Размеры, пропорции, конфигурация черепа напоминают рептилий-капторхинид. Как у рептилий, в стопе появляется астрагал. Конечности большие, хорошо развитые. В отличие от других микрозавров челюсть крепится к заднему краю черепа.

Тудитан (311-307 млн.лет, Огайо) - обратите внимание на необычные кончики пальцев.
Семейство остодолепид (290-268 млн. лет, Германия, Техас, Оклахома) - самое крупное из микрозавров, аж до 60 сантиметров, наземное, предположительно, роющее. Удлиненное туловище с маленькими мощными конечностями и укороченным хвостом. Череп клиновидный, рыло заостренное

Пелодосотис (279-272 млн.лет, Техас)

Микраротер (279-272 млн.лет, Оклахома, Техас)
Семейство пантилид (318-268 млн.лет. Канада, Нью-Мексико, Техас, Чехия) объединяет два,три или четыре рода.

Пантил (279-272 млн.лет, Техас) - небольшое (20-25 см) наземное животное, похожее на ящерицу. Огромный череп с сильными челюстями и множеством тупых зубов позволял ему охотиться на всевозможных беспозвоночных. Хвост и ноги короткие.
Семейство гимнартрид (318-268 млн.лет, Канада, Чехия, Иллинойс, Техас, Оклахома) - удлиненные с короткими конечностями. Череп маленький с одним рядом крупных конических зубов по краям челюсти. Некоторые полагают их водными формами, другие - роющими

Кардиоцефал (290-268 млн.лет, Оклахома, Техас). Как и в случае с микраротером, воду нарисовали, видимо, и-за найденных в речных отложениях единичных экземпляров.
Семейство гониорнихид монотипическое

Ринхонкос (279-272 млн.лет Оклахома) - гипотетический предок червяг. Исследователи обнаружили много сходных черт с эоцецилией - червягой юрского периода. Другие ссылаются на то, что эти черты примитивные или конвергентные.
монотипическое семейство тригекатонид (314-307 млн. лет, Колорадо) - ящерицеподобное существо вроде тудитан.
О первых тетраподах - 1, 2, 3, 4
Темноспондилы - 1, 2, 3, 4
Чешские лепоспондилы позднего карбона от Зденека Буриана. Справа на камешке темноспондил Branchiosaurus salamandroides, слева - Phlegethontia longissimum (Aistopoda), вверху - Sauropleura scalaris (Nectridea), внизу - Microbranchius pelicani (Microsauria)
Облегчение позвоночника означает, что мы больше не удерживаем тело на весу - а значит, не ходим, а ползаем, роем или плаваем. Лапы нам при таком подходе не особо нужны - и они редуцируются, вплоть до полного исчезновения. У плавающих форм могут сохраняться развитые задние лапки, у роющих - передние.
Вес тела при этом должен быть невелик - и самый крупный лепоспондил достигает длины лишь в метр. Зато очень маленьких существ может быть ОЧЕНЬ много. Поэтому, несмотря на плохую сохраняемость мелких тонких косточек, останки лепоспондил в заметных количествах находят в Северной Америке, Северной Африке и Европе.
Поздний карбон, образование Пангеи и оледенение. Европа и Северная Америка - в центре
Филогенетическое положение лепоспондил по сей день остается неясным. Из полусотни крупных таксонов позвоночных только они и анапсиды (к которым относятся черепахи) - темна вода в облацех.
Во-первых, нам совершенно неизвестны предковые виды лепоспондил - они обнаруживаются в карбоне as is уже готовыми, специализированными. А в девоне мы имеем только переходные формы, общие для всех четвероногих.
Во-вторых, отследить их потомков и братьев непросто, поскольку любое сходство может быть конвергентным. То есть, признаки совпадают не потому, что позднейшее существо происходит от примитивной амфибии, а просто в силу сходного образа жизни и решения одних и тех же задач.
Ювенильная и взрослая формы сеймурии
В-третьих, земноводные в своем индивидуальном развитии принимают две совершенно разные формы - водную ювенильную ("головастик") и половозрелую. И лепоспондилы, упростившись, педоморфны (сохраняют признаки детской формы). Не зная настоящей взрослой формы животного, как делать выводы об их систематическом положении?
Поролепиформные рыбы
Исследование кистеперых рыб показало, что тонкопозвонковые напоминают поролепиформных рыб, а не остеолепиоморфных, от которых произошли тетраподы. Получается, что лепоспондилы могут быть совершенно отдельной ветвью эволюции наземных позвоночных. Однако, оппоненты говорят, что тонкий позвоночник вполне мог возникать независимо вследствие педоморфоза. Тогда лепоспондилы могут оказаться полифилетической (не имеющей единого предка) группой. Общепринятая точка зрения - что они все-таки близкие родственники рептилиоморф и имеют одного общего предка.
Угольные леса.
Ранее некоторые считали лепоспондил предками только саламандр. С объединением лягушек, саламандр и червяг в подкласс беспанцирных от этой точки зрения отказались. Лепоспондил стали считать предками всех современных амфибий. Принятая сейчас "темноспондильная" гипотеза отказывает тонкопозвонковым в наследниках, полагая их тупиковой ветвью. Впрочем, появление доказательств отдаленности червяг от общего тренда развития голых земноводных придало убедительности гипотезе, что они таки происходят от лепоспондил, а вот все остальные - нет.
Филогенетическое древо лепоспондильной гипотезы
Традиционно базальной формой лепоспондил считались микрозавры - "крошечные ящерицы", с коротким туловищем и маленькими лапками. Добрый дедушка Кэрролл (написавший трехтомник о эволюции позвоночных) разделил их на два подотряда - тудитаноморф и микробрахоморф. Различить подотряды трудно, да, в общем, уже и не нужно. Современные кладисты очень любят менять положение групп туда-сюда и пуристически уничтожать их за грех парафилии, но вот написать толковое описание - этого от них хрен дождешься. Поэтому приходится пользоваться древней классификацией (вот тут лежит гроссбух Кэрролла и Гаскелл по микрозаврам на аглицком)
Подотряд микробрахоморф - мелкие и очень мелкие позвоночные с костяными пластинами на коже, напоминающими чешую рептилий. Микробрахоморфы практически упразднены современной систематикой, поскольку в их состав входят базальные формы микрозавров и довольно специализированные, далеко отстоящие от них брахистелехиды. Череп треугольный, шире чем у тудитаноморф, отверстия лишь для ноздрей и больших глазниц. Внутри черепа хорошо развитая "ушная капсула" - возможно здесь находилось внутреннее ухо. Морда короткая, глаза сбоку. Малое количество простых конических зубов, несколько могут быть увеличенными (напоминающими клыки), напоминает современных земноводных. Позвоночник гибкий и слабый, у пермских видов становится короче и прочнее.
Ютагерпетон (318-314 миллионов лет, Юта) - базальная форма лепоспондил, отдельные товарищи даже не причисляют их поэтому к микрозаврам
Гилоплезион (311-307 миллионов лет, Чехия) - задние лапы длиннее передних, насчитывающих лишь три пальца. Образ жизни неясен, мог быть как водным, так и наземным
Одонтерпетон (311-307 млн. лет, Огайо) - найдена только передняя половина скелета. Предположительно продолговатое тело 15 см длиной, три или четыре пальца на передних лапах.
Микробрахис (311-307 миллионов лет, Чехия) - саламандроподобное существо 15 сантиметров длиной, по видимому плавал как рыба, латеральными движениями тела и питался планктоном.
В перми семейство брахистелехид с тремя родами - батропеты (4 вида, Германия), монотипичные кэрролла (Техас, 290-279 млн лет) и квазицецилия (псевдочервяга, Техас, 279-272 млн лет)
Батропет (299-290 млн лет, Германия) - примерно 8 см длиной, череп короткий с большими орбитами.
Подотряд тудитаноморфы объединяет 7 семейств (из них два - монотипические). Вероятно у них не было водной личиночной стадии, либо она была краткой. Если верно первое, то их нельзя считать амфибиями и это параллельная поытка "амниотизации" - полностью наземной формы позвоночных. Лепоспондильный позвоночник не мог нести вес тела и спинные отростки разрастались продольно, налегая друг на друга и принимая нагрузку на себя. Хвост не приспособлен для плаванья, на нем нет нижних отростков. Конечности короткие, у большинства расположены по бокам в нижней части туловища, позволяли опираться на них и передвигаться по суше. Кожа многих несла костяные чешуи, обычно они формировали костяной панцирь с брюшной стороны, чтобы защитить ее от повреждений, но иногда были и на спине.
Форма черепа треугольная, морда короткая, заглазничная часть длиннее морды. Крупные, круглые глазницы ориентированы вперед и в стороны, что дает бинокулярное зрение в переднем направлении. Окон в черепе для выхода жевательных мышц не было, поэтому он сильно расширен в щечной области, чтобы поместить всю жевательную мускулатуру под крышей. У некоторых более поздних форм в боковой поверхности появилась щечная врезка. Зубы простые конические, у некоторых мелкие, у других - крупные, с более тупыми вершинами. Имелись небные зубы, иногда часто рассеяные по небу, иногда формирующие параллельный ряд зубов в верхней челюсти. Все это позволяло пережевывать хитин насекомых.
Семейство Гапсидопарейонтид (318-272 млн лет, Оклахома, Чехия, Германия, Канада) имело большую щечную врезку, где могла размещаться сильная жевательная мускулатура
Ллистроф (290-268 млн лет, Оклахома) - самый крупный представитель семейства, не более 20 см. Ноги короткие, но хорошо сформированные
Саксонерпетон (299-295 млн. лет, Саксония) - отсутствие щечной врезки и возможное наличие органа боковой линии намекает на то, что либо это базальная форма, либо вообще не гапсидопарейонтид.
Семейство тудитан (311-303 млн. лет, Огайо, Чехия, Канада) напоминало ящериц среднего размера. И не только внешне. Размеры, пропорции, конфигурация черепа напоминают рептилий-капторхинид. Как у рептилий, в стопе появляется астрагал. Конечности большие, хорошо развитые. В отличие от других микрозавров челюсть крепится к заднему краю черепа.
Тудитан (311-307 млн.лет, Огайо) - обратите внимание на необычные кончики пальцев.
Семейство остодолепид (290-268 млн. лет, Германия, Техас, Оклахома) - самое крупное из микрозавров, аж до 60 сантиметров, наземное, предположительно, роющее. Удлиненное туловище с маленькими мощными конечностями и укороченным хвостом. Череп клиновидный, рыло заостренное
Пелодосотис (279-272 млн.лет, Техас)
Микраротер (279-272 млн.лет, Оклахома, Техас)
Семейство пантилид (318-268 млн.лет. Канада, Нью-Мексико, Техас, Чехия) объединяет два,три или четыре рода.
Пантил (279-272 млн.лет, Техас) - небольшое (20-25 см) наземное животное, похожее на ящерицу. Огромный череп с сильными челюстями и множеством тупых зубов позволял ему охотиться на всевозможных беспозвоночных. Хвост и ноги короткие.
Семейство гимнартрид (318-268 млн.лет, Канада, Чехия, Иллинойс, Техас, Оклахома) - удлиненные с короткими конечностями. Череп маленький с одним рядом крупных конических зубов по краям челюсти. Некоторые полагают их водными формами, другие - роющими
Кардиоцефал (290-268 млн.лет, Оклахома, Техас). Как и в случае с микраротером, воду нарисовали, видимо, и-за найденных в речных отложениях единичных экземпляров.
Семейство гониорнихид монотипическое
Ринхонкос (279-272 млн.лет Оклахома) - гипотетический предок червяг. Исследователи обнаружили много сходных черт с эоцецилией - червягой юрского периода. Другие ссылаются на то, что эти черты примитивные или конвергентные.
монотипическое семейство тригекатонид (314-307 млн. лет, Колорадо) - ящерицеподобное существо вроде тудитан.
О первых тетраподах - 1, 2, 3, 4
Темноспондилы - 1, 2, 3, 4