Альтернативные стратегии поведения и индивидуальность особей

Большие полынные тетерева на току
В продолжение темы индивидуальности у животных
Я много писал про альтернативные стратегии поведения (или «жизни в целом»), когда одну и ту же проблему разные особи в популяции решают противоположными способами. Например, самцы снежного барана Ovis canadensis сanadensis на с.-з. Монтаны демонстрируют 3 стратегии спаривания[1] - основную («ухаживание», 1) и 2 альтернативные («курсирование», 2 и «блокирование», 3). В случае 1 самец просто следует за самкой в эструсе, не пробуя направлять её движений, а лишь охраняя от прочих самцов. Коитус частый и короткий (0.90±0.11 и 2.30±0.12), во время которого самка стоит. Эта стратегия свойственна самцам из верхней трети иерархии доминирования, и демонстрируется в пределах ограниченного участка территории стада, одного и того же в разные годы, чем аналогична таковой гаремных (территориальных) самцов благородного оленя.

Снежный баран
В случае 2 самец завязывает поединок с «ухаживающим» самцом и при успехе спаривается с охранявшейся им самкой (она при этом двигается). Коитус редкий (0.39±0.06 раз/час), и самки здесь с почти равной частотой спариваются с «охраняющими» и «курсирующими» самцами. Эта стратегия напоминает рейдеров благородных оленей, но осуществляется не «потихоньку», а «на глазах у всех». В случае 3 самец заранее, ещё до наступления эструса, отгоняет самку или группу самок к себе на участок, и там блокирует, а дальше действует по стратегии 1. Ряд самцов может менять стратегии в зависимости от социального контекста (и, таким образом, иметь несколько стратегий спаривания в репертуаре), но чёткость их выделения остаётся неизменной.
Альтернативные стратегии как источник социального полиморфизма
Альтернативы вроде вышеописанных лежат в основе внутривидового социального полиморфизма - когда наблюдается несколько форм отношения к территории, или взаимоотношения полов, или систем доминирования, переходящих друг в друга при изменении обстоятельств контекста, таких как плотность и интенсивность взаимодействий, корм, хищники и другие условия существования[2].
Красивый пример альтернативных стратегий, баланс которых сопрягается с динамикой популяций и изменяется в зависимости от стадий последней - поведение особей красно-серой полёвки, относящихся к цветовой морфе «чёрный огузок» (см.М.А.Потапов и др., 2011). С этим же признаком сопряжена более интенсивная окраска чепрака, см.рис.1-2. В период депрессии численности вида под Новосибирском такие зверьки встречались единично, но в период подъёма их доля быстро росла, достигнув 10-15% на пике. Аналогичные цветовые формы со сходной динамикой встречаемости отмечены и в других популяциях вида.


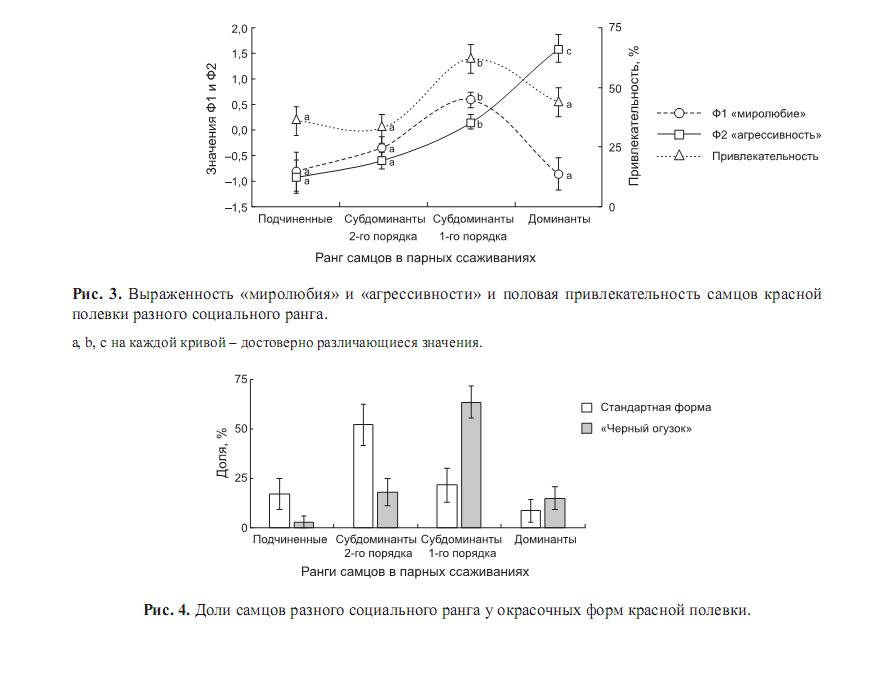
Дальше этологические эксперименты О.Ф.Потаповой с соавт. (2014) показали, что «чёрный огузок» имеет ряд преимуществ над типично окрашенными зверьками в условиях высокой плотности. Прежде всего, как положено меланистам, он «разумней дозирует» агрессию, более миролюбив в «непонятных» и новых ситуациях, и лучше отличает их от требующих агрессии, проявляет там больше исследовательской, чем тревожно-оборонительной реакции. Поэтому именно при высокой плотности такие самцы более привлекательны для самок, а также набирают большую массу и занимают больший ранг в социальной иерархии.
Вообще, у этого вида и у серой полёвки самыми привлекательными для самок оказываются субдоминанты 1-го порядка, с максимальным миролюбием при достаточной агрессивности. Частота же миролюбивых контактов партнёров при парном ссаживании - важный предиктор успешного размножения пары. Именно они преобладают среди «чёрных огузков», среди «типичных» - менее привлекательные субдоминанты 2-го порядка (доминанты же, с наибольшей агрессивностью и минимальным миролюбием, наименее привлекательны). См.рис.3-4.
Соответственно весной они оставляют больше потомков (относительно «типичных»), тем более что и начинают размножаться раньше. А к осени их доля падает, и из-за большей элиминации, и потому что приплод «типичных» поступает в популяцию позже.
Также показано дифференцированное участие разных морф в циркуляции очага клещевого энцефалита, существующего под Новосибирском. Иммунореактивность «миролюбивых» самцов существенно выше, а их, при любом ранге, больше среди «чёрных огузков». В разреженной популяции все соответствующие преимущества теряют значение, и доля «чёрных огузков» падает.
Другой пример: самцы тетерева для привлечения самок и спаривания могут собираться на токах (и очень привязаны - много лет - к выбранному току), а могут и токовать одиночно. Или самцы дрофы Otis tarda из 9 популяций, изученных в венгерской степи, в 2-х придерживаются строгой моногамии, в 7 других в разных сочетаниях наблюдаются моногамия, полигиния и промискуитет.
В конце зимы стаи дроф распадаются и самцы занимают токовые площадки. У моногамных и полигинных самцов они охраняются, развито специфическое территориальное поведение, у промискуитетных же - нет, они токуют группами рядом. Моногамные самцы спариваются лишь с одной самкой из привлечённых к месту тока, а потом постоянно держатся в первых десятках метров от гнезда. Участки полигинных самцов крупнее, охраняются лучше, и привлекают группы самок - до 4-х, между которыми устанавливается иерархия. После спаривания поведение полигинных самцов неустойчиво - он может держаться у гнезда одной из самок, как моногамный, или плюнуть на родительские обязанности и примкнуть к группам неразмножающихся самцов, как делают промискуитетные после всех спариваний. При групповом токе контакт полов ограничивается лишь краткой копуляцией. См. статьи I.Sterbetz (1980: c.32-141) и Carranza J. et al. (1989).
Читать далее