За пределами роста-3

Окончание. См.начало и продолжение.
Аннотация. Завершающая часть анализа модели пределов роста и её сопоставления с реальной динамикой. Описаны принципы моделирования и структура модели. Анализируется кибернетический механизм развития экологического кризиса, на его основе показано, почему в рамках сценария «бизнес как всегда» обычные регуляторы - рынок и технологии - не просто включаются с запозданием, но, включившись, ухудшают ситуацию. Описаны системные преимущества плановой экономики по переходу к экологически устойчивому развитию.

Птенец альбатроса, погибший от пластикового мусора
Достоинства метода моделирования
Моделирование будущего развития сложных систем ставит два важных вопроса. Первый: как моделировать это будущее, чтобы модель была состоятельна, ведь мы не пророки, а само развитие в силу нелинейности управляющих им зависимостей контринтуитивно - раз за разом расходится с выводами линейной экстраполяции, которую «по умолчанию» рвутся применять везде и всегда[1]? Второй - в чём состоит форрестеровский метод системно-динамического моделирования, чем отличается от обычных математических методов моделирования поведения сложных систем?
На первый дан общий ответ когнитивистами, занимающимися отработкой навыков управления судном, самолётом и пр. на тренажёрах, имитирующих реальное управление. При моделировании сложных систем для задач управления важна не натуралистичность, но верное отражение немногих параметров, критически важных для формируемого навыка. Надо моделировать на тренажёре и отрабатывать у людей технику управления, обеспечивающую уровень гарантированности результата выше некоторого (чем больше, тем лучше) при всём многообразии случайностей, непредсказуемых или «возмущающих» событий, возможных в реальности[2].
Здесь действует принцип - чем выше разнообразие событий контекста, независимо от которых (или даже вопреки им) мы достигаем устойчиво нужного результата[3], тем лучше. Вероятность его реализации сочетанным действием причин, учтённых в модели (и наша уверенность в его наступлении с определённой вероятностью, belief) отражает состоятельность прогноза. Видно, что данное требование к управлению системами (и, переходя в нашу область, к прогнозу развития сложной системы в будущем) означает знание - и использование - закона функционирования этих последних. Ведь закон есть регулярная, раз за разом повторяющаяся связь между переменными[4] (здесь - состояниями среды обитания под воздействием экономического развития), способная «пробить» себе дорогу в хаосе разнообразных случайностей. Совокупность последних образует контекст, в нашем случае - человеческий, или социальный.
Хороший пример - навык управления движением супертанкеров. «В начале 1980-х гг. фирма ESSO построила на озере недалеко от Гренобля учебный танкер и целую серию причальных сооружений в 1/25 реальной величины. Однако при этом время реализации команды, то есть лаг-период от подачи команды изменить курс до фактического изменения курса судна также оказывается в несколько раз меньше (в реальности он порядка 20 мин.). Поэтому тренировка вызывала отрицательный перенос - успешность проведения реальных танкеров после таких тренажёров оказывалась ниже, чем без тренировки[5]»
Важный вывод отсюда: никакая модель не может, не сможет, и не должна воспроизводить моделируемую систему в подробностях, излишняя точность здесь будет помехой. Что действительно важно - это понять, уметь выделить и отразить в модели существенные параметры, изменения которых (естественные[6] или связанные с управляющими воздействиями) критически значимы с точки зрения нашей задачи. В нашем случае это - совместима ли мировая динамика, развёртывающаяся по «стандартному сценарию», с экологической ёмкостью биосферы? Позволяют ли регуляторы, присущие «бизнесу как всегда» - рынок и технологии - вовремя «тормознуть», не допустив выхода за пределы и/или за время пребывания на кризисной траектории («за пределами роста») решить проблемы, возникшие и разрастающиеся в связи с «выходом», тем самым сделав пределы несущественными[7]? Прогоны модели на оба вопроса отвечают «нет», а авторский анализ объясняет, почему.
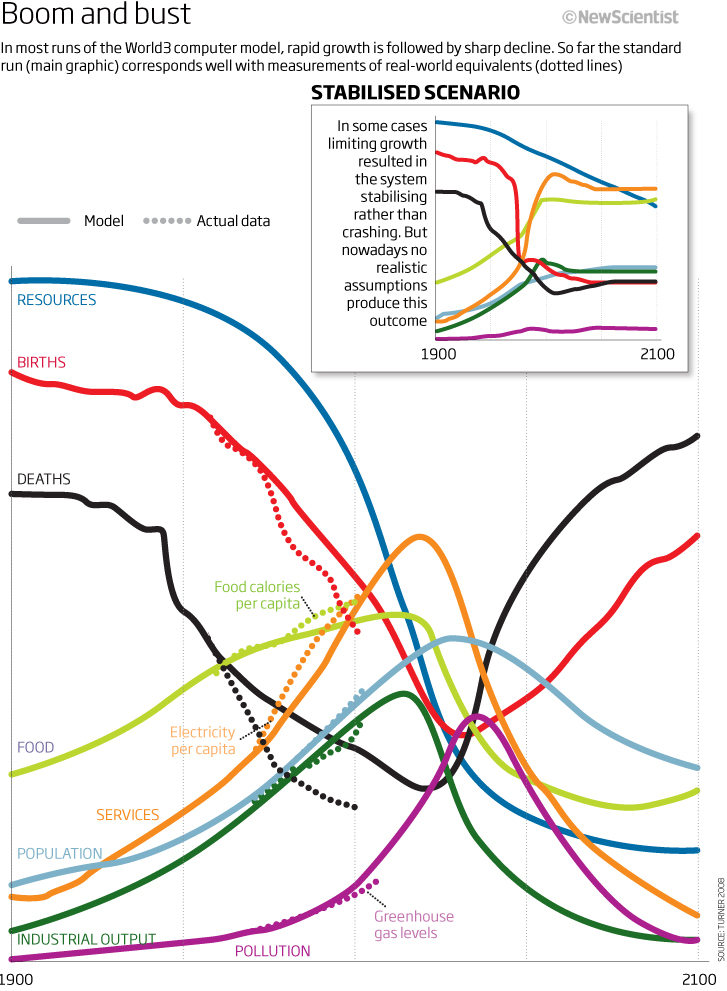
Сравнение реальных тенденций развития мира (точки) с прогнозами при стандартном сценарии (сплошные линии) и сценарии «нулевого роста» (врезка)
«Действительно, в исходную модель… не включали отдельные контуры, описывающие технологический прогресс, который мог бы автоматически решить все проблемы, связанные с экспоненциальным ростом экологической нагрузки. Это было сделано намеренно, поскольку мы и тогда не верили, и сейчас не верим, что такие технологические прорывы могут возникнуть сами собой или в результате саморегуляции свободного рынка. Развитие технологий может быть впечатляющим и даже достаточным, но только если будут приняты определённые решения в социальной сфере и если будут желаниям и средства воплотить их в жизнь. И даже если всё сложится именно так, всё равно необходимые технологии будут появляться с существенным запаздыванием. Таков наш взгляд на мир сегодня, и таким же он был 30 лет назад. Именно этот взгляд нашёл отражение в модели World3» (С.226-227).
Использованный Медоузами метод системно-динамического моделирования хорош тем, что в «каше» колебаний и трендов, составляющих мировую динамику, «отделяет тверди от вод», то есть устойчивые тенденции, обусловленные причинно-следственными связями, их переплетением и взаимодействием, от неопределённости, связанной с избыточными факторами и непрогнозируемыми событиями вроде войн или научных открытий. В чём его отличие от «обычного» математического моделирования сложных систем? Если модель состоятельна, то тенденции, прогнозируемые для будущего устойчиво реализуются в нём независимо от событий, которые модель по понятным причинам не может и не пытается прогнозировать, вроде новых научных открытий, изобретения новых технологий. Пробьют себе дорогу, как естественнонаучные законы (или почти как).
В самом худшем случае не предусмотренные моделью события и процессы воздействуют лишь количественно - сократят долю дисперсии, объяснимой моделью (скажем, с 80% до 50%), но не отменят эти последние. Иными словами, если модель состоятельна, прогнозируемые ею тенденции развития инвариантны, несмотря на неустранимые «возмущения» и новизну и устойчивы к неопределённости, которую можно задать большей или меньшей. Что есть главное достоинство метода.
При обычных методах математического моделирования разного рода процессов, развивающихся во времени, используются формулы, ставящие моделируемые характеристики в зависимость от времени или параметров других процессов. Главное, что эта зависимость постоянна на всём интервале прогноза, а не переопределяется год от года в зависимости от изменившегося состояния системы и взаимных соотношений её компонентов.
Соответственно, время здесь течёт непрерывно. Напротив, при системнодинамическом моделировании время дискретно, а формулы для расчёта (прогноза) состояний системы и её компонентов рекуррентны. Взаимные соотношения и зависимости между системными единицами и процессами[8] каждый год[9] переопределяются заново в зависимости от изменившегося состояния системы. Здесь постоянны не моделирующие зависимости, но правила их изменения, и начальное состояние, «от которого» как от печки, идут эти последние.
Так, в компьютерной популяции World3-91 индивиды ежегодно «выбирают» работу в разных отраслях экономики - добыче минерального сырья, производстве промышленных товаров, в производстве пищи, в очистке загрязнений от остальных отраслей. Или, будучи не работниками, а предпринимателями, «выбирают» те же самые отрасли для инвестирования капиталов. Затем заработанные деньги и приросшие инвестиции (вложенный капитал+полученная прибыль) вновь вкладываются в данные отрасли через потребление или реинвестиции. Так что деньги, заработанные по результатам прошлого года, начинают «работать» в следующем году.
Важно подчеркнуть: здесь моделируется чисто рыночная экономика с индивидами - homo economicus, абсолютно рациональными участниками рынка, как в роли работников/потребителей, так и в роли предпринимателей/инвесторов. Они работают и вкладывают капитал в первую очередь в тех отраслях, которые гарантируют максимальный доход, в виде зарплаты или прибыли с капитала. Не успевшие там «занять места» распределяются по другим отраслям сообразно доходности.
Поэтому по ходу развития при изменениях прибыльности разных отраслей капитал и рабочие руки направленно «перетекают» между ними, соответственно усиливая или ослабляя производственную активность. Потребительская активность индивидов однотипно меняется вслед за сдвигом соотношений «цена-качество» выпускаемых ими товаров. Поэтому линии мировой динамики не плавные, но содержат резкие скачки и изломы, в моменты, когда переопределяется знак зависимостей между элементами системы по результатам предшествующего развития.
Например, при низком уровне индустриализации экономики и урбанизации оба процесса существенно стимулируют рост населения, но при переходе некого предела начинают его ингибировать, как видим на графиках и стандартного сценария, и сценария «нулевого роста». Первый отличается от второго лишь тем, что промышленный рост в период стимуляции демографического роста в первом случае много меньше, чем во втором, а в период ингибирования - наоборот, больше. Поэтому сопоставимый подъём загрязнения от промышленного роста в стандартном сценарии «обрушивает» систему, а в «нулевом» - стабилизирует её[10], ибо до этого опережающее развитие городской и промышленной инфраструктуры «сбило» демографический взрыв, и больше прибыли от развития можно направить на купирование разного рода экологических рисков, а не на «поддержание штанов». В стандартном сценарии так не получится - взаимодействие промышленного и демографического роста с самого начала организовано так, что стимулирует демографический взрыв и потребительское давление настолько сильно, так что когда надо купировать экологические последствия этого, нельзя выделить необходимую долю капитала, почти весь он уходит на обслуживание витальных потребностей и поддержку возникших мощностей.
Параллель с логистической моделью роста популяции
Для нас, биологов, важно, что модель пределов роста по структуре и предсказаниям изоморфна логистической модели роста популяций Ферхюльста-Пирла dN/dt = r*N(1-N/K[11]). N здесь - текущая численность, r - мальтузианский параметр, мгновенная скорость роста численности, равная разности рождаемости b и смертности d в каждой точке кривой. K - предельная ёмкость среды, также определённая через численность[12]. Согласно ей, популяция S-образно растёт со скоростью r и, начиная с некоторых N, постепенно тормозит около предела, заданного ёмкостью среды (К). «Физический» смысл здесь тот же, что у движения крупного тела в вязкой среде. У долгоживущих видов, вроде копытных, рост проскакивает предел «по инерции», и если нет хищников, сокращающих численность, популяция дальше мрёт от голода и болезней, как олени на плато Кайбаб.
Важный момент состоит в том, что у подавляющего большинства видов животных зане включаются механизмы внутрипопуляционной регуляции, зависимые от плотности. В результате кривая «тормозит» сильно раньше пределов, особенно у т.н. «социальных» видов. Существенная часть ресурсов, «добытых» индивидами из среды обитания, здесь «конвертируется» не в новых особей, но в поддержание социальной структуры группировок и «привычного» уровня социальной связанности и социальной зависимости. Без этого зверьки «неспособны» результативно конкурировать, делиться на «лучших» и «худших», с поощрением первых естественным отбором и пр.
В отличие от «несоциальных» видов, у «социальных» чрезмерное разрежение так же гибельно, как и переуплотнение. Поэтому механизмы внутрипопуляционной регуляции, здесь включаются не только увеличением, но и уменьшением плотности, - например, после истребления и иной неизбирательной гибели, чтобы «собрать» уцелевших в поселения с должной плотностью социальной среды и интенсивностью контактов. Недонаселение здесь губительно также, как и перенаселение. С ростом численности таких видов частота и интенсивность контактов особей также растут[13], делая задачи регуляции всё более сложными, насущными и времязатратными, что тормозит рост.
Признаком всего перечисленного служит отмеченная у подавляющего большинства видов отрицательная корреляция между такими параметрами уравнения Ферхюльста-Пирла, как достигнутая численность N и мальтузианский параметр r. Единственное исключение - «несоциальные» виды с максимально лабильной стратегией, вроде норвежского лемминга Lemmus lemmus и азиатского бурундука Eutamias sibiricus; тут все ресурсы, добытые индивидами из среды, «конвертируются» в новых особей.
Человек отличается двумя особенностями. Во-первых, корреляция r и N в уравнении у него положительна. В отличие от животных, его популяции растут численно не за счёт большей размножаемости индивидов, а за счёт увеличения среднеожидаемой продолжительности жизни всей популяции (СОПЖ), то есть плодов общественного и научно-технического прогресса индивидам.
«С точки зрения зоолога человек отличается от всех прочих млекопитающих невозможным сочетанием двух черт популяционной динамики. Обычно зоологи делят виды на r-стратегов и K-стратегов: первые обладают высокой рождаемостью и высокой смертностью, живут, как правило, в нестабильных условиях, быстро размножаются в благоприятной ситуации, а при наступлении неблагоприятной массово гибнут. Это, например, лемминги, полёвки и многие другие мелкие грызуны и насекомоядные. К-виды обычно живут в стабильных условиях, имеют более или менее постоянную численность, медленно размножаются, могут избегать массовой гибели, но если таковая происходит - медленно восстанавливаются после неё.
Человек же - единственный биологический вид, сочетающий в себе r-тип динамики численности популяции, с К-типом её воспроизводства. Первое - это быстрые взлёты численности населения тех или иных территорий, связанные с прогрессивной урбанизацией, развитием территории и обвальные падения вследствие войн, голода, болезней и т.д., особенно в доиндустриальную эпоху.
Второе же - долгая беременность, малое количество рождений за жизнь, долгий период беспомощности детёнышей и т.д. У других видов млекопитающих, даже имеющих меньшую массу, и, значит, в потенциале - более быстрое размножение, тип популяционной динамики всегда гармонирует с типом воспроизводства, у человека - находится в контрапункте с ним.
Второй аспект, по которому Homo sapiens отличается от всех прочих видов млекопитающих, тем, что увеличение местных популяций людей всегда идёт не за счёт увеличения размножаемости индивидов, но её падения - но при увеличении средней ожидаемой продолжительности жизни, позволяющей большему числу людей в популяции до этого самого воспроизводства дожить и в нём долговременно участвовать.
Наибольшая индивидуальная размножаемость - 16-18 детей за жизнь - наблюдается в племенах австралийских аборигенов, с трудом восстанавливающих свою, в среднем постоянную численность, после периодических засух. Когда же численность населения некоторой территории долговременно (в исторических масштабах времени) растёт, число рождений за жизнь неуклонно падает при возрастании СОПЖ. Что и наблюдалось на всём протяжении истории нашей цивилизации: всякий быстрый рост населения на некоторых территориях, обычно связанный с урбанизацией и индустриализацией, сопровождается столь же быстрым падением размножаемости индивидов при опережающем росте СОПЖ, что сейчас, что в средневековье, что в эллинистическую эпоху[14].
Иными словами, только у людей воспроизводство популяции является «общим делом» в том смысле, что существующий паттерн социальной структуры, присущие ей отношения, вносят в увеличение СОПЖ существенно больший вклад, чем биологический потенциал индивидов. Не говоря уже о том, что без включения в систему социальных отношений родившиеся организмы не становятся людьми, а социальные отношения изменяются только кооперативно, в борьбе одних групп (классов, слоёв) с другими.
И наоборот - общественный регресс, «возобновляющий» действие тех факторов смертности, которые казались преодолёнными в предыдущем развитии и поэтому уменьшающий СОПЖ, ведёт не к компенсаторному росту рождаемости, но к её общему падению. Так, рост сверхсмертности в трудоспособном возрасте, вызванный увеличением социального стресса и других средовых рисков (травмы и отравления, заболевания, войны, техногенные катастрофы и ДТП) стал главным фактором снижения СОПЖ после 1991 г. во всех бывших республиках СССР и бывших соцстранах. За счёт чего как шагреневая кожа, постоянно сжималась база людей, могущих начать репродукцию, что и послужило причиной падения общего числа рождении в каждый следующий момент времени.
Соответственно, родиться ребёнок может только от тех родителей, которых минула смертность до репродуктивного возраста благодаря увеличению СОПЖ. Поэтому демографы, использующие софизм: мол, умереть не может тот, кто не родился, все умирающие сперва рождаются, поэтому подъём рождаемости наиболее важная задача. ставят телегу перед лошадью. На деле же уровень смертности в родительском поколении, обусловленной специфическими средовыми рисками данного общества, от «социальных язв» и болезней до экологии и производственных травм - причинный фактор, объясняющее, а рождаемость поколения «детей» - объясняемое, отклик. См.подробней анализ книги «Репродуктивные проблемы демографического развития России» и «Демографический переход: шаг первый, шаг последний»
Благодаря этому к размножению допускаются те, кто в более отсталых обществах вообще не дожил бы до него, не мог бы создать семью из-за бедности, чей репродуктивный потенциал был бы снижен болезнями[15] и пр. Поэтому, чтобы прийти к успеху даже в чисто дарвиновском смысле, людям надо вкладываться не в собственное размножение, а в эти плоды, развивающие общественную инфраструктуру, обслуживающую всех (пусть при неравном доступе в классовом обществе). Или как минимум в улучшение собственной позиции в обществе, но не в максимизацию размножения. Что мы и видим в демографии богатых семей в Швеции и др. странах.
Во-вторых, у нас, грешных, потребности «размножаются» сильно быстрей, чем сами люди, почему потребительское давление на среду обитания[16] несравнимо сильнее демографического. Соответственно, у животных преобразованность среды обитания предопределяется численностью и активностью индивидов: тут типичен пример бобра, зубра и странствующего голубя. У нашего вида она определяется производством, потреблением (включая рекреацию) и складированием отходов, то есть больше общественной практикой и обслуживающей её техникой, чем индивидуальной активностью. Если бы люди вторично перерабатывали бóльшую долю отходов, чем сейчас, или меньше размещали их в экосистемах, они могли бы отчуждать в свою пользу много больший процент первичной продукции, чем сейчас[17].
Соответственно, предел в модели Медоузов по смыслу тот же, что в модели Ферхюльста-Пирла - экологическая ёмкость ландшафтов[18], эксплуатируемых для нужд мирового хозяйства. Это последнее образует среду обитания человечества, аналогичную социальной среде в сообществах млекопитающих и птиц, и развивает её в ходе общественного и научно-технического прогресса. Однако предел связан не столько с численностью, сколько с потребительским давлением, разным у разных частей человечества и чем дальше, тем больше зависит не от первого, а от второго. Поэтому по оси ординат отложена не достигнутая численность N, а качество жизни разных членов популяции, оцениваемое индексом развития человеческого потенциала (Human Development Index), о котором рассказано выше.
В третьих, рост популяции животных в окрестности пределов управляется простеньким контуром из одной положительной и двух отрицательных обратных связей. Первая - прогрессия размножения, задающая темпы роста и силу «инерции» последнего. Вторые - контроль со стороны хищников, болезней с паразитами и бескормицы, с одной стороны, и плотностно-зависимые регуляторы, в основном социальной природы, с другой.
В понимании популяционной динамики конкурируют две концепции - стохастицизм и регуляционизм[19]. Первая предполагает, что популяция ведёт себя чисто мальтузиански - никаких внутренних регуляторов у неё нет, особи размножаются насколько позволяют ресурсы до тех пор, пока численность не превысит предел, за которым её снижают голод, хищники и болезни. Нижний предел численности, соответственно, определяется ресурсами. Вторая - что у популяции есть внутренние механизмы гомеостаза, которые «тормозят» рост численности задолго до достижения верхнего предела, но и не позволяют «упасть» вследствие неблагоприятных средовых флуктуаций, болезней или истребительных мероприятий. Есть и промежуточный взгляд, согласно которому при колебаниях численности нижний предел определяется средовыми факторами, верхний - регуляторными способностями популяционной системы.
В целом, чем более «социален» вид, тем больше значат внутренние регуляторы, равным образом в отношении верхнего и нижнего пределов численности, чем менее социален, тем больше значат внешние факторы.
Регуляция роста человеческого хозяйства (частью которого выступает рост численности населения, и не самой важной, в сравнении с производством) устроена намного сложнее. Самая «биологичная» её часть - воспроизводство населения - регулируется в принципе так же, как у животных, контуром из одной положительной и одной отрицательной обратной связи. Первый контур (хищники, болезни, бескормица) для нашего вида в целом не актуален в силу вышеописанного - локальные выходы человечества за предел в первый момент подталкивают общество совершенствовать технологии добычи ресурсов, чтобы пройти по кризисной траектории дальше при прежней эффективности ресурсопользования, а не «загоняют обратно» под линию К.
Второй контур, аналог плотностно-зависимой регуляции, на нас, в отличие от животных, действует двойственно. С одной стороны, главный фактор демографического перехода в популяции - распространение образования, рост числа лет, потраченных на обучение, с другой, он - лишь один из аспектов процесса развития. Все прочие составляющие (урбанизация, индустриализация, рост социального равенства, развитие медицины и рост доходов занятых в городской экономике) СОПЖ увеличивают, что, напротив, рост численности стимулирует. Однако начиная с 1980-х гг. первый аспект пересиливает второй, и чем дальше, тем больше. К 2040-2050 гг. демографический переход закончится даже в «третьем мире[20]».
Таким образом, в отличие от животных, в воспроизводстве населения контуры смертности и рождаемости не взаимодействуют со средой напрямую. Они «сцеплены», как в шестерёнчатой передаче, с изоморфными контурами воспроизводства разных видов капитала и регулируются ими через производство продуктов питания и промтоваров, с одной стороны, через производство услуг, в первую очередь образования и лечения, с другой, и через предложения рабочих мест с разной зарплатой, с третьей.
Другое отличие от популяционного роста у животных состоит в том, что циклы воспроизводства разных видов капитала оказывают на рост населения и друг на друга не однонаправленный эффект, а двойственный. До определённого предела они стимулируют рост, но по достижении определённого порога стимуляция скачком сменяется ингибированием, положительная обратная связь - отрицательной. Или наоборот (рис.17-18). Регулирование воспроизводства каждого компонента связано не с внешней средой, а с динамикой соседних циклов, и достигается за счёт стимулирующего или ингибирующего воздействия на коэффициенты.
Читать далее