Гипотезы успешности инвазивных видов растений
В продолжение темы
«Дискуссия о факторах, способствующих натурализации, продолжается и поныне. Изучение этой проблемы связано с адаптивными стратегиями вида, каждая из которых характеризуется соответствующим набором морфологических и биологических признаков. Детально этот вопрос освещен в работе Б. И. Головкина [1988], поэтому нам остается добавить лишь несколько гипотез, не вошедших в его монографию.
Интересна гипотеза о влиянии ареала растения на его способность к колонизации. Установлено [Smith, 1981], что виды, относящиеся к родам, распространенным в обоих полушариях, имеют большую способность к колонизации, чем виды, относящиеся к родам с более ограниченным распространением. Гипотеза о преимуществе видов, сочетающих вегетативный и генеративный способы размножения [Considine,1986], не вполне приемлема, поскольку подобных видов не так уж много. Гипотеза о конкурентном превосходстве видов, обладающих перекрестным опылением [Russell, Levin, 1988], как нашими, так и другими исследованиями [Pernes, 1987; Виноградова, 1992] не подтверждается. Широко распространено также мнение, что адвентивные виды должны иметь на семенах или плодах специальные приспособления для распространения животными, ветром или водой. Однако в Окленде (Новая Зеландия) из 615 натурализовавшихся видов 368 никаких явных приспособлений не имеют [Esler, 1988].
Чтобы экспериментально исследовать эту проблему необходимо сравнить биоморфологические признаки: или 1) адвентивного вида и активно вытесняемого им аборигенного вида из этого же рода, или 2) нескольких близкородственных видов, из которых только один-два стали инвазионными.
Примеров применения первой методологии очень мало. Практически полное вытеснение адвентивным видом аборигенного произошло лишь в двух случаях: в Центральной Европе Impatiens paroiflora выдворила I. noli-tangere, а в Австралии Cakile maritima вытеснила C. edentula. Евразиатский Myriophyllum spicatum, появившийся в Северной Америке около ста лет назад, в настоящее время в значительной степени вытеснил аборигенный M. exalbescens, однако изучение темпов роста и продуктивности обоих видов в различных экологических условиях не выявило заметных преимуществ заносного вида [Aiken, Picard, 1980].
При сравнении устойчивости в условиях полупустыни штата Юта (США) аборигенного Agropyron spicatum и интродуцированного из Евразии в начале XX века A. desertorum выявлены признаки, показывающие преимущества последнего [Руке, 1990]. У A. desertorum формируется в 70 раз больше семян (причем не все они после созревания сразу опадают, что позволяет избежать уничтожения вредителями и грызунами); всхожесть семян в 2 раза выше, выживаемость проростков также выше, чем у A. spicatum.
Совершенно редкий случай вытеснения одним заносным видом другого, ранее занесенного вида наблюдается на Британских островах [Simpson, 1990], где интродуцированная из Америки в 1966 году Elodea niittallii активно замещает E. canadensis. Опыты по выращиванию обоих видов при разных экологических условиях показали, что Е. niittallii растет вдвое энергичнее и образует более длинные боковые побеги, что приводит к угнетению E. canadensis в природных водоемах.
Применение второй методологии исследования можно найти в ряде работ. Опыты с пятью видами Xanthium в Австралии [Martin, Carnahan, 1983], два из которых распространились там очень широко, а три имеют ограниченный ареал, показали, что последние обладают более сильным врожденным и индуцированным покоем семян. Сравнение репродуктивного потенциала аборигенной на востоке США Celastrus scandens и быстро расселяющейся интродуцированной C. orbicidatus показало [Dreyer et al.,1987], что последняя имеет более высокие жизненность пыльцы и всхожесть семян.
Два южноамериканских перечных дерева Schinus molle и S.terebinthifolius были интродуцированы в Калифорнию более ста лет назад. Действительно натурализовался там только S. molle. Изучали прорастание семян обоих видов с целью определения механизма, препятствующего натурализации [Nilsen, Muller, 1980]. Различия обнаружены по степени холодостойкости, по отношению к воздействию фенольных кислот и во времени, протекающем между набуханием и прорастанием. Именно медленное прорастание семян S. terebinthifolius тормозит его возобновление, так как проростки не успевают развить корни во время коротких периодов обильного увлажнения почвы.
При детальном изучении натурализовавшихся растений Новой Зеландии были сформулированы [Esler, 1988] параметры «идеального сорняка», которые включают: экологическую многосторонность, быстрое созревание, плодовитость, эффективность диссеминации, вегетативное размножение и конкурентоспособность.
Таким образом, однозначного объяснения причин того, что чужеродные виды более обильны в регионах вторичного ареала, чем в регионах естественного ареала, до сих пор нет. В работе, посвященной этой проблеме [Hierro et al., 2005] приведено восемь основных гипотез для объяснения успеха чужеродных растений, внедряющихся в естественные сообщества. Первые четыре гипотезы можно проверить путем сравнения аборигенных и инвазионных популяций, последующие четыре можно оценить при изучении чужеродных видов только во вторичном ареале. Мы протестируем эти гипотезы, используя данные, полученные нами при изучении инвазионных видов Средней России.
Гипотеза 1.
Чужеродные виды не имеют естественных врагов, которые контролируют рост их популяций [Элтон, 1960]. Подтверждение гипотезы мы получили при исследовании Echinocystis lobata. В естественном ареале его популяции сильно поражают вредители и болезни, что обусловливает мозаичность распространения этого вида, тогда как в Средней России подобного явления не наблюдается [Виноградова, 2007]. Не отмечена и поражаемость вредителями у большинства других инвазионных видов. Инвазионный вид флоры Москвы Adenocaulon adhaerescens Maxim, повреждается вредителями слабо и лишь в конце вегетационного периода, тогда как в его естественном ареале на Дальнем Востоке из-за сильного поражения этого вида болезнями и вредителями с трудом можно собрать растения для гербария. Из видов, входящих в инвазионный компонент флоры Средней России, сильно поражается только Fraxinus pennsylvanica (и только в Москве), его вредителем является ясеневая изумрудная узкотелая златка, личинки которой вызывают усыхание деревьев. Однако подобное исключение только подтверждает правило, поскольку этот вид златки также инвазионный.
[также см. данные, что отсутствие пресса специфических фитофагов в новом месте произрастания позволяет им откликаться от отбор в пользу большей скорости роста, и кто лучше откликнется - тот главный захватчик. Вообще, отсутствие хищников и возможность не вкладываться в защиту от них сдвигает присутствующий у всех организмов трейд-офф между самосохранением и рискованным поведением в пользу последнего. А дальше каждый вид больше вкладывается в специфические для себя виды последнего - у гуппи в речках Тринидада это красочная раскраска самцов и половой отбор (как и у вилорогов в США), у растений - более интенсивный рост, когда могут съесть. В.К.]
Гипотеза 2.
У чужеродных видов быстро эволюируют генетические признаки, связанные с давлением естественного отбора в новых условиях среды. Гипотеза подтверждена нами при изучении Acorns calamus, Amaranthus albus, Ribes aureum, Aronia mitschurinii, Acernegundo, Impatiens glandidifera, I. parviflora, Epilobium adenocaidon, E. rubescens, Echinocystis lobata, Bidens frondosa, Conyza canadensis, Chamomilla suaveolens, Galinsoga parviflora, G. quadriradiata [Виноградова, 1992]. Выявлена общая для этих видов черта - все они на родине произрастают вне зоны экологического оптимума и потому обладают широкой нормой реакции.
Гипотеза 3.
Чужеродные виды приспособлены к типу и интенсивности нарушений природной флоры, которые являются новыми для аборигенных видов [Baker, 1974]. Думается, гипотеза представляет собой «более ученый» вариант предыдущей гипотезы - чужеродные виды действительно лучше приспособлены к высокому уровню «беспорядочности», поскольку имеют более широкую норму реакции генотипа.
Гипотеза 4.
Чужеродные виды используют ресурсы, неиспользуемые аборигенными видами [Элтон, 1960]. На начальном этапе внедрения на новые территории чужеродные виды действительно поселяются в открытых местообитаниях. Однако инвазионные виды именно вытесняют аборигенные виды, то есть конкурируют с ними за имеющиеся ресурсы. Эксперименты по сравнительной оценке конкурентоспособности близких видов, один из которых инвазионный, другой - аборигенный, проведены нами на видах рода Bidens [череда]. Было подготовлено шесть образцов, представляющих собой смесь из 50 семян североамериканской B.frondosa и 50 семян аборигенной B. tripartita, которые высеяли в пикировочные ящики, вкопанные в грядки. В двух ящиках была супесь, в двух - садовая земля, в двух - торф. Три ящика (с супесью, землей и торфом) через день-два усиленно поливали, в остальных трех растения росли без искусственного полива [Виноградова, Возна, 2008]. По всхожести семян B. frondosa получила преимущество лишь в четырех вариантах. В самых неблагоприятных условиях (в ящике с супесью без полива и в ящике с торфом при поливе) взошло на 30% больше семян B. tripartita. Однако, несмотря на это, во всех шести вариантах опыта B. frondosa имела преимущество по приросту в высоту и по биомассе (в 2-3 раза) и по семенной продуктивности (в 3-5 раз).
Опыт наглядно продемонстрировал большую конкурентоспособность инвазионного вида по сравнению с аборигенным (рис.8). Поэтому данная гипотеза в случае действительной инвазии, а не просто первоначального заноса вида на новую территорию, должна иметь прямо противоположную формулировку: «чужеродные инвазионные виды более конкурентоспособны, чем аборигенные».
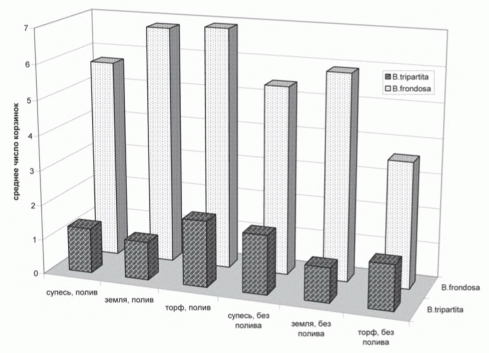
Рисунок 8. Семенная продуктивность двух видов рода Bidens на разных почвосмесях
Гипотеза 5.
Чужеродные виды могут воздействовать на виды естественных популяций биохимическими методами [Bais et al., 2003]. Гипотезу мы экспериментально не проверяли, однако полное отсутствие естественной растительности (даже тенелюбивых видов) травяного яруса в пойменных лесах из Acer negundo и Fraxinus pennsyhanica является хотя и косвенным, но вполне достаточным подтверждением гипотезы. Предварительные опыты позволили венгерским исследователям [Solymosi, 1994] предположить, что экстракт из Solidago canadensis может влиять на другие сорняки как аллелопатическое средство. Cardaria draba является активным аллелопатическим растением, ее выделения подавляют всходы других видов [Kiemnec, Mcinnis, 2002; Oasem, 2004]. Иными данными по аллелопатии видов, входящих в инвазионный компонент флоры Средней России, мы не располагаем.
[Ещё пример - завезенная в Америку из Европы в XIX веке чесночница черешковая Alliaria petiolata быстро расселяется, вытесняя местную флору. Одна из возможных причин вытеснения - выделяемые сорняком вещества препятствуют развитию симбиотических грибов, необходимых для роста местных деревьев. В.К.]
Гипотеза 6.
Чужеродные виды образуют гибриды с аборигенными, в результате чего возникает новый вид, способный к дальнейшей экспансии, а иногда и к вытеснению родительских видов. Таким образом возник, например, болотный злак Spartina х anglica - естественный гибрид афроевропейского S. maritima с американским S. alternifolia, завезенным на южное побережье Англии и закрепившимся там в начале XIX века. Сейчас S. anglica стал почти космополитом, захватив колоссальные пространства на побережье Англии, Франции, Северной и Южной Америки, Австралии и Новой Зеландии [Gufinfigou, Levasseur, 1993].
Из видов, входящих в инвазионный компонент флоры Средней России, гибридогенным является Reynoutria х bohemica - гибрид между R. japonica и R. sachalinensis. И, действительно, гибрид в экспериментальной посадке дает больший прирост по биомассе, чем родительский вид R. japonica.
В Москве высокую активность проявляет Symphytum х uplandicum, гибрид между местным S. officinale и кавказским S. asperum. Amelanchier spicata, по-видимому, также является естественным гибридом между североамериканской A. canadensis, интродуцированной в 1590 году из провинции Квебек (Канада) для королевского ботанического сада в Париже, и введенной в культуру в 1596 году A. ovalis, являющейся единственным видом ирги с европейским ареалом [Куклина, 2006а]. [Всё это примеры поддерживаемого отбором «заимствования» чужих генов через гибридизацию, ввиду большей успешности гибридов в местообитаниях гибридной зоны, о котором я писал ранее. В.К.]
Гипотеза 7.
Многовидовые сообщества более стойки к вторжению чужеродных видов, чем маловидовые [Элтон, 1960]. Теоретически увеличение видового богатства значительно снижает возможности инвазии, видимо потому, что более разнообразные сообщества полнее и эффективнее используют имеющееся пространство, ограничивая ресурсы для местных видов. Снижение биоразнообразия, таким образом, создает благоприятные условия для инвазий в эту систему, потенциально ускоряя потерю разнообразия и гомогенизацию биот на земном шаре. Эта гипотеза, разработанная на основе теоретического моделирования, впоследствии была подтверждена экспериментально в лабораторных условиях в микрокамерах с контролируемыми гомогенными условиями обитания. Однако правомерность этой гипотезы для естественных сообществ остается дискуссионной, поскольку экспериментальных данных, полученных именно в природных неконтролируемых условиях, до сих пор мало. В связи со спорностью вопроса нами был проведен эксперимент по искусственному внедрению Bidens frondosa в естественные сообщества с варьирующим числом слагающих их видов и с различными доминантами [Виноградова, 2003].
Вопреки теории оказалось, что на участках без явных доминантов и с минимальным числом видов в фитоценозе всхожесть семян B. frondosa была самой низкой и составляла 10%. Вовсе не проросли семена на участке без доминантов с максимальным числом видов в фитоценозе. Зато 100%-ная всхожесть наблюдалась на двух участках: на первом монодоминантом была Zizania aquatica и росло еще шесть видов, а на втором - монодоминантом выступала Carex rostrata и росло еще пять видов. В полидоминантных сообществах независимо от числа слагающих их видов всхожесть составляла 10-40%. Следовательно, помимо биоразнообразия фитоценозов, не последнюю роль играет и их структурированность. Сообщество противостоит натиску чужеродных видов, если состоит из 2-3 выраженных доминирующих и 6-10 сопутствующих видов. [что есть важный вклад в жерихинскую концепцию биоценотической регуляции 'эволюции. В.К.]
Гипотеза 8.
Изменчивость уровня инвазибельности различных естественных сообществ зависит от числа чужеродных видов, вторгающихся в сообщество [Williamson,1996]. Думается, эта гипотеза вполне правомерна. Во всяком случае, число адвентивных видов, отмеченных по берегам рек, обочинам дорог и на пустырях, значительно превышает их число в лесных или степных фитоценозах. Соответственно имеется прямая корреляция и между числом адвентивных и инвазионных видов. Разработка методики экспериментальной проверки этой гипотезы станет для нас очередной задачей.
Очевидно, что успешность расселения заносных видов невозможно объяснить лишь одной причиной. Процесс натурализации задействует целый комплекс факторов, и, вероятно, для разных видов определяющие факторы в этом комплексе будут различными».
Чёрная книга флоры Средней России