Эволюция кооперации и альтруизма: от бактерий до человека-2
...9 . Склонность к альтруизму сильнее у тех, кому нечего терять
Вот еще один пример, показывающий, что альтруизм общественных насекомых очень далек от идеала бескорыстности.

Осы Liostenogaster flavolineata живут семьями, включающими от 1 до 10 взрослых самок, из которых только одна - самая старая - откладывает яйца, а остальные заботятся о личинках.
Когда царица погибает, ее место занимает следующая по старшинству оса.
Внешне помощницы ничем не отличаются от царицы, однако жизнь они ведут гораздо более тяжелую и опасную: если царица почти не покидает гнезда, то помощницам приходится летать за кормом для личинок, изнашивая крылышки и рискуя попасться на глаза хищнику.
С переходом помощницы в ранг царицы ожидаемая продолжительность ее жизни резко увеличивается.
У этого вида, как и у многих других, осы-помощницы сильно различаются по степени «трудового энтузиазма». Одни, не жалея себя, проводят в поисках пищи до 90% времени, а другие предпочитают отсиживаться в безопасном гнезде и вылетают за кормом на порядок реже.
На первый взгляд кажется, что эти различия трудно объяснить с позиций теории родственного отбора, поскольку степень трудового энтузиазма помощниц не зависит от степени их родства с царицей и личинками, о которых они заботятся.
Как выяснилось, каждая помощница строго дозирует свой альтруизм в зависимости от того, насколько велики ее шансы стать царицей и оставить собственное потомство.
Если эти шансы туманны и зыбки (как у низкоранговых молодых ос, последних в «очереди» на царский престол), то имеет смысл работать поактивнее, чтобы хоть через чужих детей передать свои гены следующим поколениям. Если же помощница имеет высокий ранг, ей выгоднее поберечь себя и поменьше рисковать.
Этот вывод основан на результатах изящных экспериментов. Из одной семьи удаляли осу, занимающую второе место в иерархии (то есть первую по старшинству после царицы), а из другой, такой же по размерам семьи - удаляли низкоранговую молодую осу. После этого следили за поведением осы, до начала эксперимента занимавшей в иерархии третье место.
В первом гнезде эта оса после удаления старшей помощницы повысила свой ранг, переместившись с третьего места на второе, во втором - осталась на третьем месте. Размер обеих семей остался одинаковым.
Выяснилось, что в первом случае оса начинает работать примерно вдвое меньше.
Во втором случае, когда из гнезда изымалась низкоранговая помощница, оса номер три продолжала работать столько же, сколько и раньше (см.: Склонность к альтруизму сильнее у тех, кому нечего терять).
Эти результаты показывают, что величина «альтруистического усилия» у ос действительно регулируется в зависимости от шансов данной осы на собственный репродуктивный успех.
Появление такого поведения в ходе эволюции на самом деле хорошо объясняется «правилом Гамильтона» (просто нужно учитывать, что величина c, то есть цена альтруистического поведения, меняется в зависимости от обстоятельств, в том числе от шансов на «царский престол»).
10. Чтобы предотвратить появление обманщиков, нужно обеспечить генетическую идентичность кооператоров
Можно ли создать общественное устройство, где альтруизм будет поддерживаться без насилия, и при этом не будет никаких обманщиков и эгоистов? Ни осам, ни людям это пока не удалось. Но некоторые кооперативные симбиотические системы, существующие в природе, указывают на то, что в принципе можно не допустить самого появления обманщиков.
Для этого нужно свести генетическое разнообразие индивидуумов в кооперативной системе к полному нулю. Это исключает возможность конкуренции между генетически различающимися разновидностями симбионтов за то, кто из них более эффективно будет эксплуатировать общие ресурсы (ухватит себе больший кусок общего пирога).
Если все симбионты генетически идентичны, эгоистическая эволюция внутри системы становится невозможной, потому что из минимального набора условий, необходимых для эволюции, - дарвиновской триады «наследственность, изменчивость, отбор» - исключается один из компонентов, а именно изменчивость.
Симбионтам-близнецам не важно, кто из них ухватит для себя больший кусок, ведь с точки зрения естественного отбора они все - просто одно и то же. Их эволюционные «интересы» автоматически отождествляются с интересами всей системы. При этом отбор перестает действовать на уровне индивидуальных симбионтов и начинает действовать на уровне целых симбиотических систем.
Именно поэтому эволюции так и не удалось, несмотря на многократные «попытки», создать нормальный многоклеточный организм из генетически разнородных клеток. Все настоящие многоклеточные организмы образуются из клонов - потомков одной-единственной клетки.
Рассмотрим этот механизм на примере такой интереснейшей кооперативной симбиотической системы, как сельское хозяйство у насекомых.
Если кооперативная система состоит из крупного многоклеточного «хозяина» и маленьких «симбионтов», то для хозяина самый простой путь обеспечить генетическую идентичность симбионтов - это передавать их вертикально, то есть по наследству, причем заниматься этим должен только один из полов - либо самцы, либо самки.
Именно так передаются, например, митохондрии у всех эукариот - строго по материнской линии, причем сами митохондрии размножаются клонально. Так же передают из поколения в поколения свои сельскохозяйственные культуры муравьи-листорезы.
При вертикальной передаче генетическое разнообразие симбионтов автоматически поддерживается на близком к нулю уровне за счет генетического дрейфа и «бутылочных горлышек».
Существуют, однако, и симбиотические системы с горизонтальной передачей симбионтов. В таких системах симбионты у каждого хозяина генетически разнородны, они сохраняют способность к эгоистической эволюции, и поэтому среди них то и дело появляются обманщики.
Например, известны штаммы обманщиков среди светящихся бактерий (симбионтов рыб и кальмаров), азотфиксирующих бактерий-ризобий (симбионтов растений), микоризных грибов, зооксантелл (симбионтов кораллов).
Во всех этих случаях эволюции «не удалось» обеспечить генетическую однородность симбионтов, и поэтому хозяевам приходится бороться с обманщиками иными методами - например, иммунологическими, или просто терпеть их присутствие, полагаясь на те или иные механизмы, обеспечивающие баланс численности обманщиков и честных кооператоров - скажем, на парадокс Симпсона или на балансирующий отбор, который основан на том, что часто быть обманщиком выгодно только до тех пор, пока численность обманщиков не слишком высока (иначе будет некого обманывать).
Все это не так эффективно, но что поделаешь: естественный отбор замечает только сиюминутную выгоду и совершенно не интересуется отдаленными эволюционными перспективами.
Для того чтобы развился механизм, обеспечивающий генетическую однородность симбионтов, этот механизм должен давать немедленную выгоду, иначе отбор его не поддержит. Та выгода, о которой мы говорили до сих пор, - лишение симбионтов возможности эволюционировать в обманщиков - как раз относится к категории «отдаленных перспектив» и поэтому не может работать как эволюционный фактор на микроэволюционном уровне.
Но если уж какому-то виду так повезет, что вертикальная передача симбионтов будет для него сопряжена с какой-то сиюминутной выгодой и поэтому будет закреплена отбором, это может обеспечить его отдаленным потомкам триумфальный успех.
Термиты подсемейства Macrotermitinae, освоившие эффективное сельское хозяйство - выращивание грибов - до сих пор казались исключением из правил.
Передача симбионтов (одомашненных грибных культур) у них не вертикальная, а горизонтальная, однако грибы-обманщики в их огородах полностью отсутствуют.

Симбиоз термитов с грибами возник единожды свыше 30 млн лет назад в экваториальной Африке и оказался очень успешным.
В настоящее время подсемейство термитов-грибоводов включает 10 родов и около 330 видов, играющих важнейшую роль в круговороте веществ и функционировании тропических сообществ Старого Света.
В отличие от грибов, выращиваемых муравьями-листорезами, одомашненные термитами грибы уже утратили способность к самостоятельному существованию. Они растут только в термитниках на специально обустроенных грядках из растительного материала, пропущенного через кишечник термитов.
Основав новую колонию, термиты собирают в окрестностях споры грибов Termitomyces и засевают ими свои плантации. Естественно, исходный посевной материал оказывается генетически весьма разнородным.
Грибы образуют в термитнике особые маленькие плодовые тела (нодулы), содержащие бесполые споры (конидии). «Бесполыми» эти споры называют потому, что они образуются без мейоза, а их геном идентичен геному родительской грибницы.
Конидии служат исключительно для размножения грибов внутри термитника. Термиты питаются нодулами, а споры проходят через их кишечник неповрежденными и используются для засева новых плантаций.
Грибам нужно позаботиться и о том, чтобы попасть в новые термитники. Конидии за пределы термитника обычно не распространяются. Для этого используются половые споры (базидиоспоры). Они образуются в плодовых телах другого типа - больших, прорастающих сквозь стены термитника наружу.
Это «нормальные», обычные плодовые тела, характерные для грибов-базидиомицетов (к базидиомицетам относятся почти все съедобные грибы, плодовые тела которых мы собираем в лесу).
Из базидиоспор, принесенных термитами в новое гнездо, вырастают маленькие гаплоидные мицелии (грибницы). Клетки разных гаплоидных мицелиев сливаются и превращаются в дикарионы - клетки с двумя гаплоидными ядрами. Из них вырастают уже «настоящие», большие дикариотические мицелии, способные образовывать плодовые тела.
Слияние ядер у базидиомицетов происходит только при образовании базидиоспор, непосредственно перед мейозом. Конидии содержат по два гаплоидных ядра, как и клетки мицелия, а базидиоспоры - по одному.
Таким образом, грибы производят маленькие плодовые тела в основном для термитов («альтруизм»), а большие - в основном для себя («эгоизм»). Стратегия грибов-обманщиков могла бы состоять в том, чтобы производить побольше больших плодовых тел и тратить меньше ресурсов на кормление термитов. Но среди грибов Termitomycesобманщиков нет, и до сих пор никто не знал, почему.
Эту загадку разгадали совсем недавно. Оказалось, что в каждом термитнике выращивается только один штамм грибов. При этом в разных термитниках культивируются разные штаммы.
Таким образом, стало ясно, что термиты предотвращают появление обманщиков обычным способом - при помощи монокультурного разведения симбионтов. Но как им удается создавать монокультуру из изначально разнородного посева?
Оказалось, что всё объясняется особенностями взаимоотношений между штаммами грибов при плотном посеве - в сочетании с тем фактом, что размножение грибов внутри термитника полностью контролируется термитами. Выяснилось, что у Termitomycesсуществует положительная корреляция между частотой встречаемости штамма в смешанной культуре и эффективностью его бесполого размножения.
Иными словами, генетически идентичные мицелии помогают друг другу - но не другим мицелиям - производить конидии.
Исследователи обнаружили, что между относительной численностью штамма в смешанной культуре и эффективностью его размножения существует положительная обратная связь. Это неизбежно приводит к формированию монокультуры уже через несколько циклов «пересевов», осуществляемых термитами.
Какова природа этой положительной обратной связи? Дело в том, что отростки дикариотических мицелиев могут срастаться друг с другом, но только в том случае, если эти мицелии генетически идентичны. Чем крупнее мицелий, тем больше ресурсов он может направить на производство нодул и конидий. Это способствует росту урожайности в монокультуре и вытеснению «меньшинств».
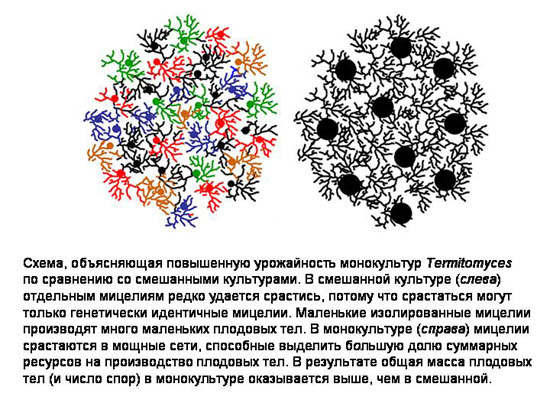
По-видимому, дикий предок грибов Termitomyces оказался удачным кандидатом для доместикации (одомашнивания) как раз потому, что был склонен образовывать монокультуры при плотном посеве.
Повышенная урожайность монокультур могла стать тем самым «сиюминутным преимуществом», которое позволило отбору поддержать и развить эту склонность на ранних этапах формирования симбиоза.
В долгосрочной (макроэволюционной) перспективе она оказалась решающей, потому что избавила термитов-грибоводов от угрозы появления грибов-обманщиков.
В конечном счете это обеспечило симбиотической системе эволюционный успех (см.: Выращивание монокультур - ключ к эффективности сельского хозяйства у термитов).
Между прочим, при переходе людей от охоты и собирательства к производству продовольствия (в ходе «неолитической революции», начавшейся 10-12 тысяч лет назад) проблема выбора кандидатов для доместикации тоже стояла крайне остро. Хороший симбионт - большая редкость, и во многих регионах просто не нашлось подходящих видов животных и растений.
Там, где их случайно оказалось больше всего, человеческая цивилизация стала развиваться с наибольшей скоростью. Об этом подробно рассказано в замечательной книге Джареда Даймонда «Ружья, микробы и сталь» (doc-файл, 2,66 Мб).
Из всего сказанного видно, что если бы не проблема обманщиков, порождаемая отсутствием у эволюции дара предвидения и заботы о «благе вида» (а не гена), наша планета, вероятно, была бы царством всеобщей любви и дружбы. Но эволюция слепа, и поэтому кооперация развивается только там, где то или иное стечение специфических обстоятельств помогает обуздать обманщиков или предотвратить их появление.
Существует не так уж много удачных «инженерных решений», позволяющих справиться с проблемой обманщиков. На каждое из этих решений эволюция неоднократно «натыкалась» в своих блужданиях по пространству возможного.
11. Межгрупповая конкуренция способствует внутригрупповой кооперации
Рассмотрим еще один механизм эволюции кооперации и альтруизма, который позволит нам перейти к рассмотрению биологического объекта, который нас традиционно интересует больше всего, а именно нас самих.
Если у какого-то вида животных кооперация уже развилась настолько, что вид перешел к общественному образу жизни, то дальше начинаются интересные вещи. Во многих случаях оказывается, что индивид может успешно размножиться только будучи членом успешной группы.
При этом конкуренция обычно существует не только между особями внутри группы, но и между группами. К чему это приводит, показывает модель вложенного перетягивания каната, которую разработали американские этологи.
Целью исследователей было найти простое объяснение четырем закономерностям, наблюдаемым в социальном устройстве общественных насекомых. Эти четыре закономерности перечислены на слайде.
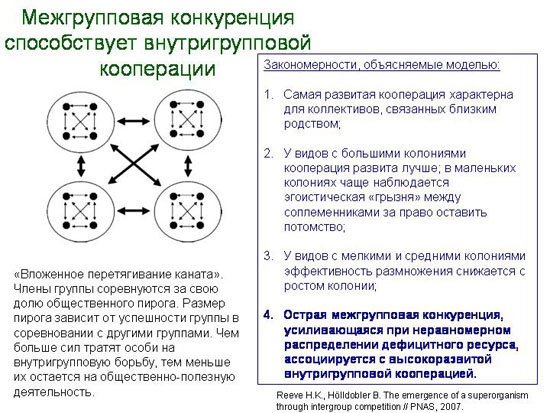
В модели «вложенного перетягивания каната» каждый индивид эгоистически расходует часть «общественного пирога» на то, чтобы увеличить свою долю этого пирога. Эта потраченная на внутригрупповые склоки часть называется «эгоистическим усилием» данного индивида.
Доля, доставшаяся в итоге каждому индивиду, зависит от соотношения его собственного «эгоистического усилия» и суммы «эгоистических усилий» остальных членов группы.
Нечто подобное наблюдается у общественных насекомых, когда они осуществляют «взаимный надзор» - мешают друг другу откладывать яйца, стараясь при этом отложить свои.
На тех же принципах строятся в модели и взаимоотношения между группами. Таким образом, получается «вложенное», двухуровневое перетягивание каната. Чем больше энергии тратят особи на внутригрупповую борьбу, тем меньше ее остается для межгруппового «перетягивания» и тем меньше получается «общий пирог» группы.
Исследование этой модели при помощи теории игр показало, что модель хорошо объясняет наблюдаемые закономерности.
Авторы вывели ряд уравнений, описывающих долю ресурсов, доставшихся в итоге каждой особи при той или иной степени ее эгоистичности, и нашли для разных ситуаций «эволюционно стабильную» величину индивидуального эгоистического усилия, то есть такую величину, при которой никакие мутации, меняющие эту величину в ту или иную сторону, не дадут преимущества своим носителям и не смогут распространиться в генофонде.
Модель показала, что внутригрупповая кооперация должна расти с ростом внутригруппового родства. Это соответствует идеям Гамильтона и Холдейна о том, что степень родства между членами группы - отнюдь не второстепенный фактор, а мощный регулятор развития кооперации.
Но модель также предсказывает, что кооперация может иметь место даже при полном отсутствии родства между членами группы. Для этого необходима острая конкуренция между группами.
Этим можно объяснить, например, странный факт из жизни пустынных муравьев Acromyrmex versicolor, у которых некоторые самки, способные основать новую колонию, отказываются от этой возможности ради помощи другим таким же самкам, совершенно неродственным - особенно в том случае, если компания самок-основательниц подвергается опасности набегов со стороны уже существующих колоний.
Главный вывод состоит в том, что межгрупповая конкуренция - один из важнейших, а может быть, и самый главный фактор, стимулирующий развитие кооперации и альтруизма у социальных организмов (см.: Межгрупповая конкуренция способствует внутригрупповой кооперации).
Теоретически эта модель может быть приложима не только к насекомым, но и к другим социальным животным, и даже к человеческому обществу.
Аналогии вполне очевидны. Ничто так не сплачивает коллектив, как совместное противостояние другим коллективам; множество внешних врагов - обязательное условие устойчивого существования тоталитарных империй и надежное средство «сплочения» населения в альтруистический муравейник.
12. Альтруизм у людей зависит не только от воспитания, но и от генов
Прежде, чем применять к человеку какие-то модели, разработанные в рамках эволюционной этики - а эволюция альтруизма представляет собой центральную тему эволюционной этики - мы должны убедиться, что человеческая нравственность имеет хотя бы отчасти наследственную, генетическую природу, что она подвержены наследственной изменчивости, и поэтому на нее может действовать отбор.
На пчелах, бактериях и других общественных организмах, не способных к культурной эволюции, изучать становление альтруизма проще, поскольку сразу можно уверенно предполагать, что разгадка кроется в генах, определяющих поведение, а не в воспитании, культуре, традициях и т. д.
С приматами, особенно с человеком, сложнее: здесь помимо обычной биологической эволюции, основанной на отборе генов, необходимо учитывать еще и социальную и культурную эволюцию, основанную на отборе идей, или мемов (в данном случае речь идет о таких мемах, как морально-нравственные нормы, правила поведения в обществе и т. п.)
Исследования последних лет показали, что моральные качества людей в значительной мере определяются генами, а не только воспитанием.
Причем необходимо подчеркнуть, что имеющиеся методы позволяют оценивать только «верхушку айсберга» - только те наследственные черты, по которым у современных людей еще сохранилась изменчивость, которые еще не зафиксировались в нашем генофонде.
Многие из аллелей, которые обеспечивали рост альтруизма у наших предков, наверняка давно уже зафиксировались, то есть достигли стопроцентной частоты. Они есть у всех людей, и поэтому такие методы, как близнецовый и сравнительно-генетический анализ, уже не могут их выявить.
Ясно, что способность к альтруистическому поведению в основе своей «заложена» в наших генах - ведь кооперация была необходима нашим предкам задолго до того, как они овладели речью и тем самым создали «питательную среду» для распространения и эволюции мемов.
Ясно, что практически любой здоровый человек при соответствующем воспитании способен научиться вести себя более или менее «кооперативно» и «альтруистично». Значит, некий генетический «базис» альтруизма есть у всех нас - соответствующие гены прочно зафиксировались в человеческой популяции.
Однако до сих пор имеется очень мало экспериментальных данных, на основании которых можно судить о том, в какой фазе находится эволюция альтруизма в современном человечестве: то ли «генетический» этап давно закончился, и сегодня актуальными являются только социально-культурные аспекты этой эволюции, то ли эволюция альтруизма продолжается и на уровне генов.
В первом случае следует ожидать, что наследственная изменчивость людей по признакам, связанным с альтруизмом, очень мала или вовсе отсутствует, а столь очевидные всем нам различия по уровню доброты и порядочности объясняются исключительно воспитанием, условиями жизни и разными случайными обстоятельствами.
Во втором случае мы должны ожидать, что эти различия отчасти объясняются также и генами. «Отчасти» - потому что роль внешних факторов в становлении человеческой личности слишком очевидна, чтобы кому-то пришло в голову ее отрицать.
Вопрос ставится следующим образом: оказывают ли индивидуальные генетические различия хоть какое-то влияние на наблюдаемую вариабельность людей по степени кооперативности, альтруизма и взаимного доверия.

В поисках ответа на этот вопрос используется, в частности, близнецовый анализ. При помощи специальных тестов определяют степень альтруизма (или, например, таких качеств, как доверчивость и благодарность) у множества пар однояйцовых и разнояйцовых близнецов, а потом сравнивают сходство результатов у разных пар.
Если однояйцовые близнецы больше похожи друг на друга по своей доброте, чем разнояйцовые, это веский довод в пользу генетической природы данного признака.
Такие исследования показали, что склонность к добрым поступкам, доверчивости и благодарности имеет в значительной мере генетическую природу и подвержена наследственной изменчивости у современных людей.
Наблюдаемые у людей различия по степени доверчивости и благодарности как минимум на 10-20% предопределены генетически (см.: Доверчивость и благодарность - наследственные признаки).
Это очень серьезный вывод, имеющий далеко идущие последствия. Он означает, что биологическая эволюция альтруизма в человечестве еще не закончена. В популяции сохранился полиморфизм по генам, определяющим большую или меньшую склонность к кооперативному поведению и взаимному доверию.
По-видимому, в разных природных, социальных и экономических условиях естественный отбор благоприятствует то доверчивым кооператорам, то недоверчивым эгоистам, и переменчивость этих условий способствует сохранению разнообразия.
Есть и другой вариант объяснения, основанный не на переменчивости условий, а на частотно-зависимом «балансирующем» отборе.
Чем больше кругом доверчивых альтруистов, тем выгоднее паразитировать на чужой доброте; но если паразитов становится много, их стратегия оказывается уже не столь выгодной, да и общество начинает воспринимать их как реальную угрозу и вырабатывает меры для обуздания эгоизма.
Выявляются и конкретные гены, влияющие на моральные качества человека. Приведу один пример. Сейчас активно изучается действие нейропептидов окситоцина и вазопрессина на социальное поведение животных и людей.
В частности, оказалось, что у людей перназальное введение окситоцина повышает доверчивость и щедрость. С другой стороны, близнецовый анализ показывает, что эти черты характера являются отчасти наследственными. Это позволило предположить, что те или иные аллели генов, связанных с окситоцином и вазопрессином, могут влиять на склонность людей к альтруистическому поведению.
Недавно генетикам удалось обнаружить связь между некоторыми аллельными вариантами гена окситоцинового рецептора (OXTR) и склонностью людей проявлять бескорыстный альтруизм. Окситоциновый рецептор - белок, вырабатываемый некоторыми клетками мозга и отвечающий за их восприимчивость к окситоцину. Аналогичные свойства обнаружены также и у гена вазопрессинового рецептора (AVPR1a).
В регуляторных областях этих генов имеются так называемые однонуклеотидные полиморфизмы (single nucleotide polimorphisms, SNP). Это такие нуклеотиды, которые могут быть разными у разных людей (большая часть каждого гена, естественно, одинакова у всех людей).
Оказалось, что некоторые из аллелей этих генов обеспечивают меньшую, а другие - большую склонность к альтруизму (см.: Найден ген, влияющий на склонность к добрым поступкам).
Все это говорит о том, что альтруизм у людей даже сегодня еще может развиваться под действием биологических механизмов, а не только социально-культурных факторов.
Расширенная версия доклада на IV Международной конференции "Биология: от молекулы до биосферы" (15.12.2009)
Александр Марков
***
Окончание следует.
Источник.
НАВЕРХ.