Первый кит бегущий краем моря

Эволюция и происхождение китообразных долгое время оставалось загадкой для палеонтологов. Из-за скудости палеонтологической летописи вопрос происхождения китов был причиной ожесточенных споров креационистов с учеными, отстаивающими эволюционное учение. Ископаемые останки, проливающие свет на развитие и становление этой удивительной группы животных до самого недавнего времени были очень редки. Несомненно, современные киты являются вторичноводными млекопитающими,- в процессе эволюции их предки сначала вышли из воды, дав начало амфибиями и рептилиями, а затем снова вернулись в воду млекопитающими. Произошло это примерно 50-55 миллионов лет назад, в позднем палеоцене-эоцене.
Хоть в это и трудно поверить, глядя на современного голубого кита, но все китообразные, включая китов, дельфинов и морских свиней, являются потомками сухопутных млекопитающих отряда парнокопытных (конечно не современных, а древних копытных).
Ранее традиционные взгляды на эволюцию китообразных состояли в том, что их ближайшими родственниками и, вероятно, предками являлись мезонихии - вымерший отряд хищных копытных, которые напоминали волков с копытцами вместо когтей и были сестринской группой парнокопытных. Эти животные имели зубы необычной конической формы, похожей на зубы китообразных. В частности из-за этого учёные долгое время считали, что китообразные произошли от некой предковой мезонихии. Однако новые молекулярно-генетические данные свидетельствуют о том, что китообразные - близкие родственники парнокопытных, в частности ныне живущих гиппопотамов. На основе этих данных сейчас предлагается даже включать китообразных в состав отряда парнокопытные и предлагается название «Cetartiodactyla» для монофилетического таксона, включающего две эти группы. Однако наибольший возраст известных ископаемых остатков антракотериев, предков бегемотов, пока на несколько миллионов лет меньше, чем возраст пакицета, наиболее древнего из известных предков китов.
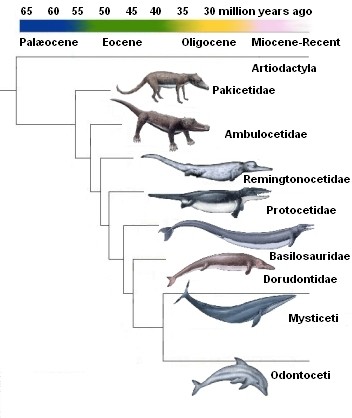
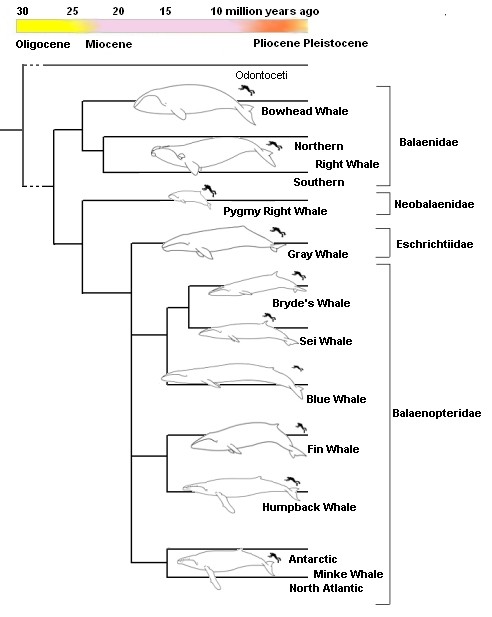
Основная схема эволюции китов
Обо всем расскажет ухо
Во время экспедиции американского палеонтолога Филиппа Джинджериша (P.Gingerish) в Пакистане, ученые получили весьма интересный материал. Они искали остатки эоценовых наземных млекопитающих в местах, где они уже находились, но им попадались только морские организмы. Около 50 млн. лет назад в этом районе проходила, меняющаяся со временем, береговая линия древнего моря Тетис, которое разделяло на протяжении большей части эоценового периода Евразию и Африку. Среди остатков рыб и моллюсков палеонтологами были найдены два фрагмента тазовых костей, явно принадлежавших относительно крупным «ходящим» животным. В то же время в другой части Пакистана была обнаружена челюсть примитивного парнокопытного.
Спустя два года в Северном Пакистане экспедицией Джинджериша была обнаружена еще одна странная находка. Это был кусок черепа странного создания размером с волка. Рядом были обнаружены остатки других млекопитающих, на этот раз наземных, живших около 50 млн. лет назад. Однако череп найденного неизвестного зверя имел черты, напоминавшие некоторые детали строения слуховой системы современных китообразных.


Пакицетус
Напомним, в водной и воздушной среде звуковые волны распространяются по-разному. У китов которые живут в наше время нет наружного уха, а слуховой проход, ведущий к среднему уху, или крайне сужен, или вообще отсутствует. Барабанная перепонка утолщена, неподвижна и не выполняет те функции, которые свойственны наземным животным. Их у китов берет на себя так называемая слуховая булла - особое костное образование, изолированное пазухами. Булла в черепе неизвестного животного, открытого Джинджеришем, хотя и не была по-настоящему «китовой» и явно не могла обеспечить хороший подводный слух, но тем не менее отличалась достаточно характерными изменениями. Выходило, что это существо - его назвали по месту находки пакицетусом (Pakicetus) - могло быть одной из первых эволюционных ступенек на пути перехода от наземных животных к китообразным. В то же время можно было предполагать, что загадочный зверь имел и обычную функциональную барабанную перепонку, позволяющую воспринимать звуки, разносящиеся по воздуху, - на земле он пока проводил никак не меньше времени, чем в воде. Строение скелета пакицета еще раз подтвердило то, что киты не являются прямыми потомками мезонихид. Напротив, предки китов отделились от парнокопытных и перешли к водному образу жизни уже после того, как сами парнокопытные отделились от общих с мезонихидами предков. Таким образом, протокитообразные были ранними формами парнокопытных, которые сохранили некоторые характерные для мезонихид признаки (коническую форму зубов), утраченные современными парнокопытными. Интересно, что самые ранние предки всех копытных млекопитающих были, вероятно, частично плотоядными или «мусорщиками», всеядными животными.
Пакицеты были копытными животными, иногда их классифицируют как ранних китов. Они жили на территории современного Пакистана (откуда и название - «кит из Пакистана») в раннем эоцене, около 50 миллионов лет назад. Это было животное, внешне напоминавшее собаку, однако с копытцами на пальцах и с длинным тонким хвостом. Первоначально предполагалось, что ухо пакицета было хорошо приспособлено для жизни под водой, однако, как показали дальнейшие исследования, уши пакицета подходят только для воздушной среды, и, если пакицет действительно предок китов, возможность слышать под водой была позднейшей адаптацией уже имевшегося слухового аппарата. Как утверждает американский палеонтолог Ханс Тевиссен, зубы пакицета уже напоминают зубы ископаемых китов.

Еще одна реконструкция пакицета - «с волосами»
Тевиссеном было также обнаружено, что схожее устройство уха наблюдалось в окаменелостях еще одного необычного существа - небольшого похожего на оленя животного индохиуса. Индохиус (буквально «свинья Индии») - маленькое (размером с кошку) существо хрупкого сложения, остатки которого найдены на территории Кашмира (Индия). Чаще всего его сравнивают с современными африканскими водяными оленьками; сходство нарушает только длинный хвост - общая черта различных групп примитивных млекопитающих раннего кайнозоя. Возраст этого существа оценивается в 48 миллионов лет. Индохиус классифицируется как представитель семейства раоеллид (Raoellidae) - примитивных парнокопытных. Его считают представителем сестринской группы по отношению к ранним китообразным на основании особенностей строения все той же ушной области. Слуховая булла индонихуса, образованная из барабанной кости также очень необычна по форме и демонстрирует особенности строения, характерные для открытых незадолго до этого древнейших китов, и, в частности, все того же пакицетуса. Это небольшое - размером с домашнюю кошку - травоядное животное обладало некоторыми чертами, сближающими его с китами и свидетельствующими об адаптации к водной среде. В их числе толстая и тяжёлая костная оболочка, напоминающая костную оболочку некоторых современных полуводных животных, таких как гиппопотамы, что способствует уменьшению плавучести и, как следствие, позволяет оставаться под водой. Это позволяет предположить, что индохиус, подобно современному водяному оленьку, нырял под воду, чтобы укрыться от хищника. Так, в его останках повышено содержание изотопа кислорода 18О, что указывает на водный образ жизни. Однако, содержание изотопа углерода 13С говорит о том, что он редко кормился в воде. Впрочем, столь же вероятно, что его пища могла состоять из высших водных растений (цветковых). В любом случае, судя по изотопному составу эмали зубов, Indohyus наверняка не входил в пищевые цепочки, в основании которых лежит пресноводный фитопланктон, образованный водорослями, а не высшими растениями.


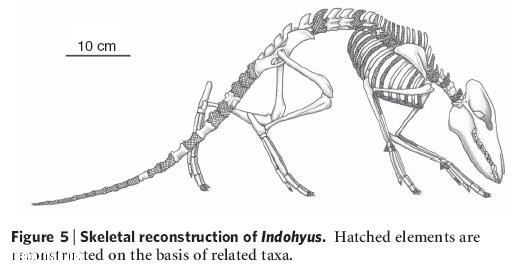
Индохиус
«Крокодил среди млекопитающих»
Наиболее примечательным из древних китов является известный многим амбулоцет, известный из эоцена Пакистана. Внешне это млекопитающее походило на трёхметрового крокодила.
«Чудовище, неподвижно лежавшее в воде среди мангровых зарослей, заметило добычу - подходящего по размерам зверя, пришедшего на водопой. За нескольких энергичных толчков задних ног оно приблизилось к берегу, запустило свои мощные зубы в тело жертвы и попятилось назад в воду. Когда крепко зажатое в челюстях животное, не имея возможности вздохнуть, перестало биться, хищник выполз на берег, чтобы начать трапезу на твердой земле. На первый взгляд чудовище походило на крокодила - с короткими ногами, массивным хвостом, длинной вытянутой мордой и высоко посаженными, выступающими над поверхностью головы глазами. Однако его тело покрывали не пластинки панциря, а шерсть, ноги заканчивались не когтями, а чем-то напоминающим копыта, а зубы - это были зубы зверя, а не пресмыкающегося…», -так в представлении палеонтологов выглядит амбулоцетус, один из первых китов.
Земля в среднем эоцене - 50 млн. лет назад
Амбулоцет был полуводным животным: его задние лапы лучше приспособлены для плавания, нежели для ходьбы по суше. Вероятно, он плавал, изгибая тело в вертикальной плоскости, как современные выдры, тюлени и киты. Предполагается, что амбулоцетиды охотились подобно современным крокодилам, подстерегая в засаде рыб и пришедших на водопой животных. В челюсти амбулоцета уже имелось начало характерного для китов канала, проводящего звук к уху. Положив нижнюю челюсть на грунт - как это делают крокодилы - амбулоцетус «лоцировал» передвижение по берегу своих потенциальных жертв.
Близкими родственниками амбулоцета были ремингтоноцетиды. Представители этого семейства были мельче по размеру, имели более удлинённую морду и были лучше приспособлены к подводной жизни. Предполагается, что образом жизни они напоминали современных выдр, охотясь из засады на рыб.
У представителей обеих групп ноздри располагались на конце морды, как у наземных млекопитающих.
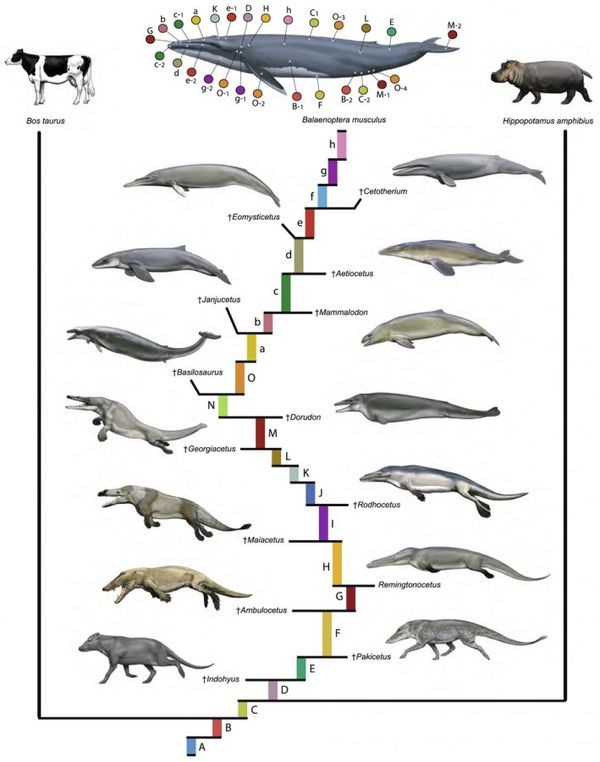
Ближайшие родственники китов сегодня - гиппопотамы
Стоит отметить важные изменения черепа в процессе эволюции китообразных - перемещение глазниц с верхнего (как у крокодилов) положения у пакицетуса и амбулоцетуса на бока головы, как у протоцетид и современных китов. Ноздри сместились с вершины морды у пакицетуса на вершину головы (дыхало) у современных китов. Зубы стали простыми и однообразными - приспособленными только для удержания, а не разжевывания добычи. У усатых же китов они исчезли совсем; их «китовый ус» - роговые пластины, с зубами никак не связаны.
Анализ изотопного состава атомов кислорода, присутствующих в зубах ископаемых китов, позволяет делать заключения о том, жили ли они в пресной или морской воде - во второй присутствует большая доля изотопа 18О. Получилось, что организм пакицетусов получал только пресную воду, амбулоцетусы могли жить как в пресных, так и в соленых водоемах, а протоцетиды были уже настоящими морскими животными.


Амбулоцет. Четко видна «крокодилообразная» форма скелета
«Протокиты»
Протоцетиды образуют крупную и разнообразную группу, известную по находкам в Азии, Европе, Африке и Северной Америке. Данное семейство включает в себя большое количество родов, некоторые из них довольно хорошо изучены (например, родоцетуса, известный из третичных отложений Белуджистана). Все известные протоцетиды обладали хорошо развитыми передними и задними конечностями, которые могли поддерживать тело на земле; вероятно, они вели амфибиотический образ жизни, обитая как в водной среде, так и на суше. Пока не ясно, имелся ли у протоцетид хвостовой плавник, как у современных китообразных, однако очевидно, что они были неплохо приспособлены к водному образу жизни. Например, крестец - часть позвоночника, к которой крепится таз - у родоцета состоял из пяти раздельных позвонков, в то время как позвонки в крестце наземных млекопитающих слиты. Носовые отверстия сдвинулись у протоцетид еще дальше вверх по рылу - это является первым шагом к расположенным на макушке ноздрям нынешних китообразных. Версия об амфибийной природе протоцетид подкреплена находкой беременной самки майацета сокаменевшим плодом, повёрнутым головой к выходному отверстию. Это заставляет предположить, что роды у майацета проходили на суше - в противном случае детёныш имел шансы захлебнуться.


Кутхицетус
О происхождении ранних китов от копытных говорят такие особенности, как, например, наличие копыт на концах пальцев у родоцета. У данного китообразного кости нижнего отдела передней конечности были сжаты и уже напоминали ласты, а длинные нежные ступни, возможно были перепончатыми. Связки между позвонками, формирующими крестец, у родоцетуса были ослаблены, что позволяло позвоночнику изгибаться так, чтобы создавать волнообразные вертикальные движения хвоста. По мнению Джинджериша, он плавал «по-собачьи» на поверхности, а под водой передвигался за счет совмещения толчков веслообразных задних ног и хвоста. Скорее всего, этот зверь еще не полностью порвал с наземной средой и периодически выходил на сушу, где передвигался толчками, - наподобие современных ушастых тюленей. Вообще, в течение эоцена китообразные совершили резкий скачок в морфологических изменениях: из четвероногих наземных животных они превратились в полностью водные формы, совершенно не похожие внешне на свои наземных предков и родственников. Возможная причина этого явления - отсутствие конкурентов в новой среде обитания.

Родоцет
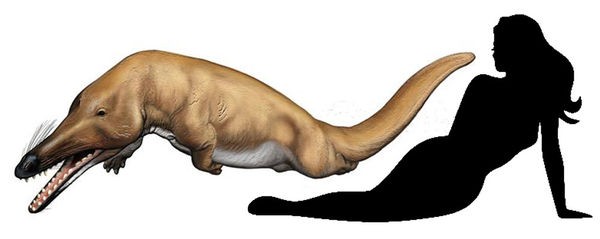
Ремингтоноцет
Выход в океан
От протоцетид, произошли уже вполне «дельфинообразные» дорудоны (Dorudon) - возможные предки базилозавров и современных китов, постепенно расселившихся по всем морям земного шара.
Базилозавр (обнаруженный в 1840 г . и первоначально принятый за рептилию, чем объясняется «рептильное» имя) и дорудон жили приблизительно 38 миллионов лет назад и представляли собой уже чисто морских животных. Базилозавр был столь же велик, как крупные современные киты, достигая порой 18 метров в длину. Дорудонтиды были несколько меньше, до 5 метров .
В связи с переходом к чисто водному образу жизни у базилозаврид наблюдается деградация задних конечностей - они, хотя и хорошо сформированы, невелики и уже не могут использоваться для передвижения. Впрочем, возможно, они играли вспомогательную роль при спаривании. Тазовые кости базилозаврид уже не связаны с позвоночником, как это было у протоцетид.

Джорджияцет
Как у современных китов, плечо у дорудонов и базилозавров оставалось подвижным, а локоть и запястье образовывали передний плавник. Однако вопрос о том, когда именно киты окончательно потеряли задние конечности, остается открытым. Например, у настоящего усатого кита, чьи остатки были совсем недавно обнаружены в слоях возрастом 27 млн. лет, еще имелись неплохо сформированные ноги.
В хвостовом отделе дорудона имелся позвонок округлой формы, подобный тому, который имеется у современных китов в основании хвостового плавника. Так что, возможно, дорудоны и базилозавры уже имели вполне китовый хвостовой плавник.


Дорудон
Между тем, эти киты еще не были «настоящими китами». Луо (Zhe-Xi Luo) - палеонтолог, сотрудник Музея естественной истории в Питтсбурге, показал, что у базилозавров и дорудонтов - первых полностью водных китов - слуховая система по строению была уже достаточно близка к слуховой системе современных китов. Однако при всём сходстве с современными китами у базилозаврид и дорудонтид отсутствовал лобно-жировой выступ, так называемая дыня, позволяющая ныне существующим китообразным эффективно использовать эхолокацию. Мозг базилозаврид был сравнительно небольшого размера, из чего можно предположить, что они вели одиночный образ жизни и не имели такой сложной социальной структуры, как у некоторых современных китообразных.


Базилозавр
Возникновение «китового уса»
Китовый ус уникален для усатых китов, но зубатые киты, не имея его, тем не менее, также являются китами. Поэтому данный признак нельзя считать каким-то основополагающим: это частная адаптация одной группы китообразных. На протяжении следующего за эоценом олигоценового периода уровень моря понизился. «Прото-Индия» соединилась с Азией (результатом этого «столкновения» стало возникновение Гималаев), а Австралия и Антарктида удалились друг от друга, в результате чего образовалось широкое свободное кольцо морей в Южном полушарии. Возникло южное циркумполярное течение, начал образовываться ледяной панцирь. Это создавало новые условия для живущих в морях млекопитающих, что, по мнению ряда специалистов, и привело к возникновению современных подотрядов - усатых и зубатых китов. Наиболее древней известной переходной формой между ними и древними археоцетами является Llanocetus, первичный усатый кит, найденный в антарктических отложениях возрастом около 34 млн. лет. Судя по всему, он вполне мог питаться крилем. Зубатые же киты, по мнению специалистов, возникли примерно в то же время, развивая способности к эхолокации, которая позволяла активно охотиться в глубине.

Выход на сушу и выход в океан
К сожалению, находки остатков первых представителей двух современных отрядов крайне редки. Понижение уровня моря в олигоцене осушило прибрежные районы, которые могли содержать эти остатки, и они подверглись разрушению. Но раскопки в более поздних слоях показывают, что немного времени спустя, 30 млн. лет назад, настоящие усатые и зубатые киты были представлены несколькими семействами.
Три года назад, в 2011 году учеными были найдены окаменелые останки одного из древнейших усатых китов, которые оказались «недостающим звеном» в эволюции китового уса. Исследователи обнаружили, что огромные эластичные челюсти синих китов и их собратьев развились из более жёстких образований.
На верхних челюстях усатых китов располагаются несколько сотен роговых пластин, которые работают как фильтр, отсеивающий планктон из поступающей в ротовую полость животного воды. Киты набирают в рот очень большое количество воды, широко раскрывая пасть за счет того, что у них нет жесткого соединения между двумя половинами нижней челюсти. Кроме того, практически все китообразные обладают очень широким черепом, что дополнительно увеличивает возможный максимальный объем попадающей в рот воды. Благодаря своему способу питания киты смогли эволюционировать до столь внушительных размеров.


Janjucetus hunderi - один из первых усатых китов
Ученые точно не знают, какой была последовательность возникновения этих двух характерных особенностей - большого черепа и гибкого сочленения челюстей. Авторы новой работы описали кости древнего китообразного «Janjucetus hunderi», обитавшего в земных океанах около 25 миллионов лет назад, и новые данные свидетельствуют в пользу гипотезы о том, что изначально у китов сформировался широкий череп.
«Ранние усатые киты были лишены одного из признаков всех живых (и большинства ископаемых) усатых китов - свободного сустава нижней челюсти, - отмечает автор исследования Эрих Фицджеральд из Музея Виктории (Австралия). - Без него нынешние усатые киты просто не смогли бы питаться так, как они привыкли».
Учёный имеет в виду следующее: кит распахивает нижнюю челюсть под очень большим углом; эластичная ткань, прикреплённая к челюсти, растягивается, и это позволяет животному набирать в рот огромный объём воды. Пластины из китового уса, растущие из верхней челюсти, выполняют роль своеобразного сита, отфильтровывающего криль - основной источник питания.
Новые останки принадлежали виду «Janjucetus hunderi», который жил около 25 млн лет назад у берегов Австралии и имел, вероятно, около трёх метров в длину, то есть был размером со среднего дельфина. Его украшали большие зубы для захвата и измельчения добычи, чем он сильно отличался от сегодняшних усатых китов с их волосовидными зубами. К тому же, как уже было сказано, его нижняя челюсть не могла распахиваться так широко.
Тем не менее «J. Hunderi» был усатым китом, ибо обладал рядом адаптаций, характерных для этого подотряда. Например, это широкая верхняя челюсть, свидетельствующая о том, что большой рот появился прежде способности к фильтрации. Две половины нижней челюсти «J. Hunderi» были прочно соединены между собой и не позволяли древнему морскому млекопитающему очень широко открывать пасть. При этом верхние челюсти животного выглядели типично для современных китообразных, а сам череп был очень широким. Ученые полагают, что «J. Hunderi» не фильтровал поступающую воду, проталкивая ее в обратную сторону изо рта, а заглатывал вместе с оказавшейся там добычей.
Среди современных крупных китов кашалот даёт, как ни странно, возможность ответить на вопрос как киты перешли к фильтрующему типу питания. Обычно в популярной литературе кашалота изображают закусывающим гигантским кальмаром. Но это, хотя и известная, но далеко не столь часто встречающаяся добыча. Естественно, на китобойных станциях при разделке кашалотов из их желудков извлекали таких моллюсков. Но также известно, что часто в желудках кашалотов находили большое количество сравнительно мелких кальмаров и рыб. Даже если рыба достигала порой метровой длины, она всё равно мала по сравнению с многотонным кашалотом. Современные зубатые киты часто питаются мелкими рыбами и кальмарами, просто засасывая их ртом. У клюворылых китов значительно редуцированы зубы - иногда всего лишь до двух крупных зубов, явно не приспособленных для схватывания такой добычи, как рыба и кальмары.

Голубой кит не имеет зубов
У самых ранних усатых китов («усатых» скорее по анатомии скелета, а не по наличию китового уса) зубы достаточно редко сидят на челюстях. Можно предположить, что они постепенно утрачивали функцию захвата и непосредственного удержания добычи, и служили скорее для «запирания» рта. Кстати, у современной китовой акулы мелкие зубы выполняют именно такую роль. Первоначальная добыча прото-усатых китов, скорее всего, была достаточно крупной, а мелкая рыба имела возможность проскользнуть между зубами хищника. Не стоит улыбаться: современная сельдь тоже имеет реальный шанс выскользнуть даже из гораздо более совершенного фильтровального аппарата кита-горбача. Собственно говоря, так ловит добычу, мелких плавающих ракообразных, современный тюлень-крабоед (Lobodon), у которого зубы приобрели специфическую форму, став многовершинными и плоскими.
Канадского исследователя Эдварда Д. Митчелла удивило необычное строение зубов кита Llanocetus: они сидели в челюстях с большими промежутками, имели неглубокие корни и глубокие выемки на коронке, делящие зуб на лопасти. Популярная статья, в которой рассказывалось про открытие этого животного, называлась »Ancient whale smiled like a sieve» («Улыбка древнего кита похожа на решето»). Это наглядное подтверждение того, что первые усатые киты использовали зубы, чтобы закрывать выход из своего рта мелким кормовым объектам, которых схватывали по несколько штук. В процессе эволюции у китов развивалось приспособление, которое одновременно позволяло воде свободно выходить из пасти животного, но более эффективно задерживало мелкую рыбу, ракообразных или кальмаров, оказавшихся во рту кита. Ключ к познанию дальнейшего изменения ловчего аппарата китообразных также даёт знание об «испорченных» генах, отвечающих за развитие зубов, найденных в геноме современных китов. При дальнейшем развитии китового уса наличие зубов постепенно стало нейтральным признаком, и мутанты с «испорченными» генами, отвечающими за развитие зубов, не отличались по успешности выживания от китов с неповреждёнными генами, у которых отрастали анатомически полноценные зубы. В дальнейшем исчезновение зубов сняло анатомические ограничения на развитие более совершенного цедильного аппарата.

Llanocetus
Эхолокация
Настоящие зубатые киты (Odontocetes) осуществляют эхолокацию, создавая серию щелчков на различных частотах. Звуковые импульсы излучаются посредством лобной дыни, отражаются от объекта и регистрируются с помощью нижней челюсти.
Эхолокация - система ориентации в пространстве по задержке возвращений отраженной звуковой волны - появилась у предка нынешних дельфинов и зубатых китов более 28 миллионов лет назад. Адъюнкт-профессор Ньюйоркского технологического института Джонатан Гайслер (Jonathan Geisler) провелисследование ископаемого вида «Cotylocara macei», открытого в окрестностях Чарльстона (Южная Каролина). «Самые важные результаты нашего исследования касаются эволюции эхолокации и сложной анатомии, обеспечивающей эту способность. Она возникла примерно в ту же эпоху, когда происходила диверсификация китов - разные размеры тела и мозга, разные способы питания», - говорит Гайслер.
Зубатые киты, дельфины и морские свиньи издают высокочастотные звуки сквозь закрытую область в носовых каналах, за дыхалом - тогда как у всех остальных млекопитающих (в том числе людей) звуки рождаются в гортани. У зубатых китов механизм очень сложен - это множество мускулов, воздушных полостей и жировых прослоек, втиснутых в небольшой участок лица.Палеонтологи считают, что такая сложная система развивалась постепенно, шаг за шагом.


Cotylocara macei - один из первых китов с развитой эхолокацией
По мнению Гайслера, кит «Cotylocara macei» был способен к эхолокации. «Плотные кости и воздушные пазухи его черепа помогали сфокусировать его звуки в единый поток звука, который позволял киту искать себе пищу ночью или в мутной воде», - говорит ученый.
Проведя сопоставительный анализ, Гайслер и его коллеги определили, что Cotylocara принадлежал к вымершему семейству китов, отделившихся от остальных китообразных как минимум 32 миллиона лет назад. По всей видимости, рудиментарная форма эхолокации возникла у общего предка «Cotylocara» и других зубатых китов примерно 35-32 миллиона лет назад.
«Cotylocara» отличались рядом уникальных анатомических черт, в том числе глубокой полостью на верху головы (отсюда и название вида - «голова с пазухой»), где при погружении в воду животные «запасали» воздух - и та же полость, вероятно, отражала звуки, идущие со стороны лица. Также обращает на себя внимание похожая на радиолокационную антенну кость вокруг носовых отверстий, которая также могла отражать звук и улучшать качество эхолокации. «Анатомия черепа очень необычна. Ничего такого я не видел ни в одном ките - ни живом, ни ископаемом», - говорит Гайслер.
Изучение черепов сквалодона («Squalodon») позволяет предположить первичное возникновение эхолокации и у этого вида китов. Сквалодон жил с начала среднего олигоцена до середины миоцена, около 33-14 миллионов лет назад, и имел ряд признаков, сходных с современными зубатыми китами. Так, например, сильно сплюснутый череп и вынесенные челюстные дуги наиболее характерны для современных Odontoceti. Несмотря на это, возможность происхождения современных дельфинов от сквалодона считается маловероятной, хотя сквалодон дает представление об эволюции китов на ранней стадии.

Сквалодон
В данном посте я не буду подробно рассматривать вопрос появления и развития эхолокации у зубатых китов, иначе придется писать тут целый научный труд и рассмотреть множество точек зрения по этому вопросу. В целом, можносказать о том, что последние ископаемые останки китообразных еще раз блестяще подтвердили верность дарвинизма - эволюционного учения. Будущие открытия докажут верность этого тезиса со всей очевидностью, как на примере эволюции китов, так и других групп живых организмов.

Постепенное смещение у китов носовых пазух к затылку
Интересно то, что в ноябре 2006 года, близ берегов Японии был пойман живым дельфин афалина с недоразвитыми, но хорошо заметными снаружи задними конечностями. Его фото обошло, наверное, все ленты новостей. Этот атавизм лучше всего свидетельствует о том, что предки китов жили на суше.

Современная реконструкция левиафана, гигантского зубатого кита, у него эхолокация уже также была развита:



[источники]
источники
http://www.infuture.ru/article/10699
http://www.pro-kitov.info/history.php
http://www.nat-geo.ru/article/637-kityi-v-pustyine/
http://www.sivatherium.narod.ru/
Вот пожалуйста вам еще про Ужас древних морей , а так же Версия: Как погибали динозавры. А вот место, где Идут скелеты строем … и говорят даже, что Бронтозавры не вымерли ?
Оригинал статьи находится на сайте ИнфоГлаз.рф Ссылка на статью, с которой сделана эта копия - http://infoglaz.ru/?p=51980